
Novophytoptus luzulis, Chetverikov, Philipp E., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4006.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:D01B5A31-9D9E-4C50-9443-101EDB3E04C3 |
|
DOI |
https://doi.org/10.5281/zenodo.6103077 |
|
persistent identifier |
https://treatment.plazi.org/id/03B39162-FF9C-0C61-E9D1-3679BE70FE08 |
|
treatment provided by |
Plazi |
|
scientific name |
Novophytoptus luzulis |
| status |
sp. nov. |
Novophytoptus luzulis n. sp.
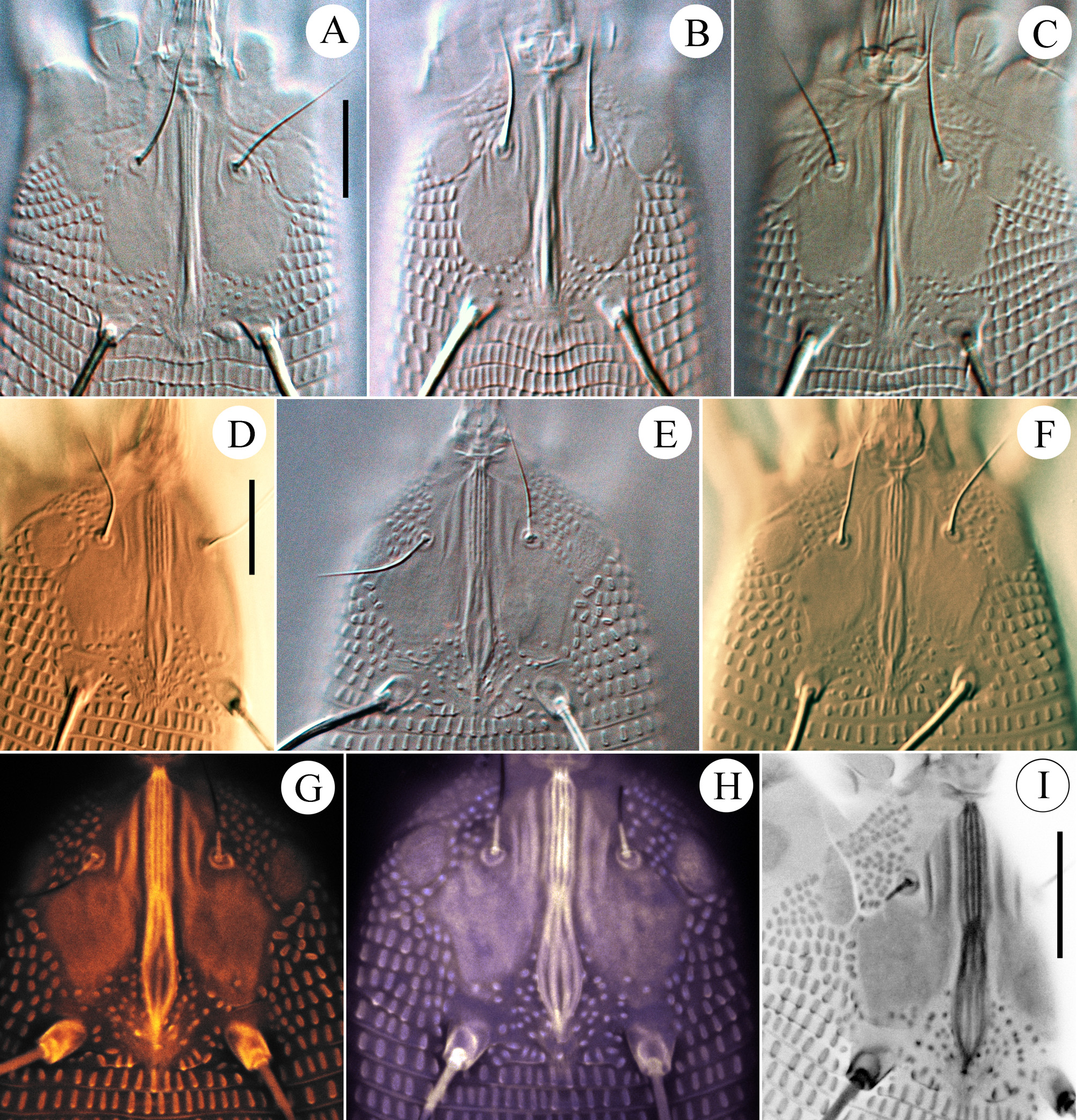
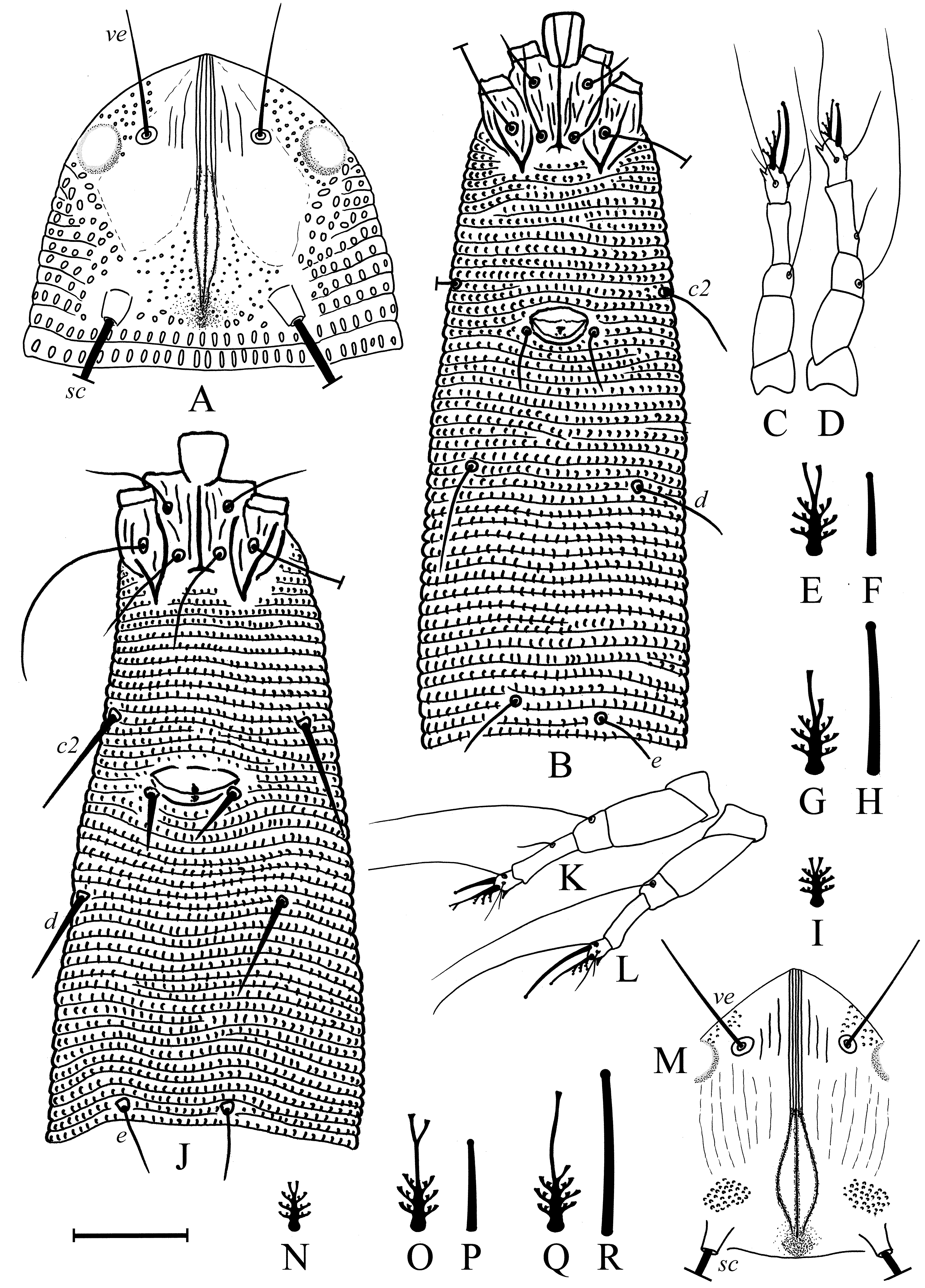
( Figs. 1 View FIGURE 1 D–I, 2 D–F, 3 D–F, 4, 5 A–I, 6 A–C, 7 E–F, 8)
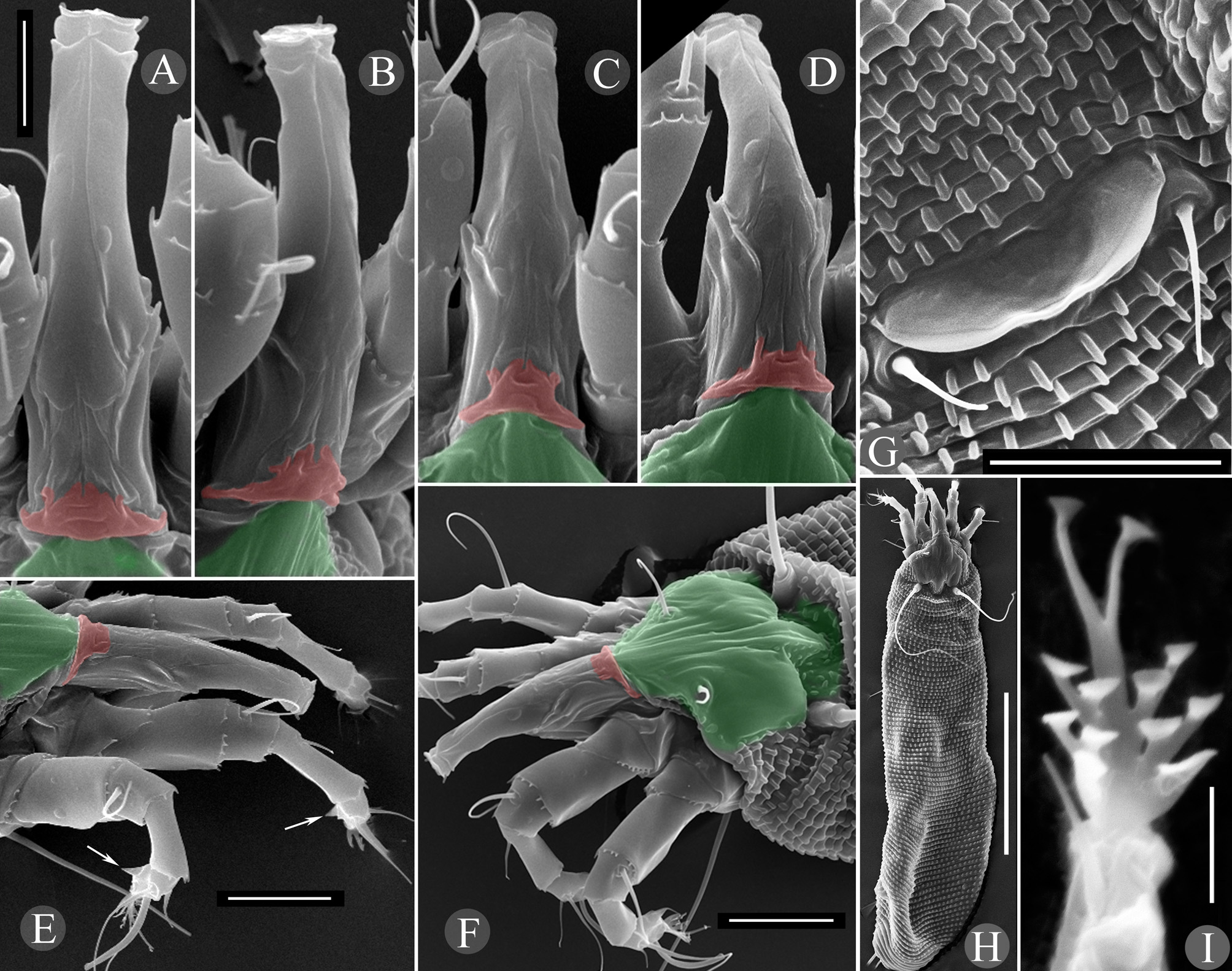
Female (n=9: holotype & paratypes). Idiosoma vermiform, 341 (270–356), 54 (42–54) wide at the level of setae c2, 68 (45–68) wide at the level of setae f. Prodorsal shield ( Fig. 1 View FIGURE 1 D–I) subpentagonal, 33 (27–34) x 25 (24–28) wide with two ocella-like spots lateral to setae ve. Anterior margin of prodorsal shield forming a triangular extremity resting on enlarged fused basal parts of chelicerae and associated with basal gnathosomal structures of unknown origin ( Fig. 4 View FIGURE 4 , coloured pink). Shield ornamentation with several median longitudinal ridges, tightly packed, forming a very narrow figure of 8 or bottle-shape with a “neck” centrally (better seen in Fig. 1 View FIGURE 1 H,I). Anteriorly, this “bottle” includes five thin ridges, whereas posteriorly, only three (rarely four) are observed (see Fig. 1 View FIGURE 1 H). The ridges converge anteriorly and posteriorly. Two to three thin lines present between tubercles ve and anterior part of “bottle”. Rudimentary posteromedian fovea (“pit” sensu Keifer) surrounded by small scattered lines on rear prodorsal shield margin; 57 (52–80) circular/oval granules between sc tubercles. Setae ve 16 (11–19), directed anterolaterad, tubercles 15 (12–15) apart; sc 109 (74–116) long, directed posterolaterad, tubercles 15 (12– 15) apart. Distance between tubercles ve–sc 20 (16–22). Gnathosoma elongate, directed forward and slightly ventrad, 23 (21–25); pedipalp coxal seta ep 6 (3–6), pedipalp genual seta d absent ( Fig. 4 View FIGURE 4 ), subapical pedipalp tarsal seta ν 0.5 (0.5–1). Ventrally, basal gnathosoma covered by petal-like suboral plate bearing two cuticular ridges, forming V-shaped figure ( Fig. 3 View FIGURE 3 E).
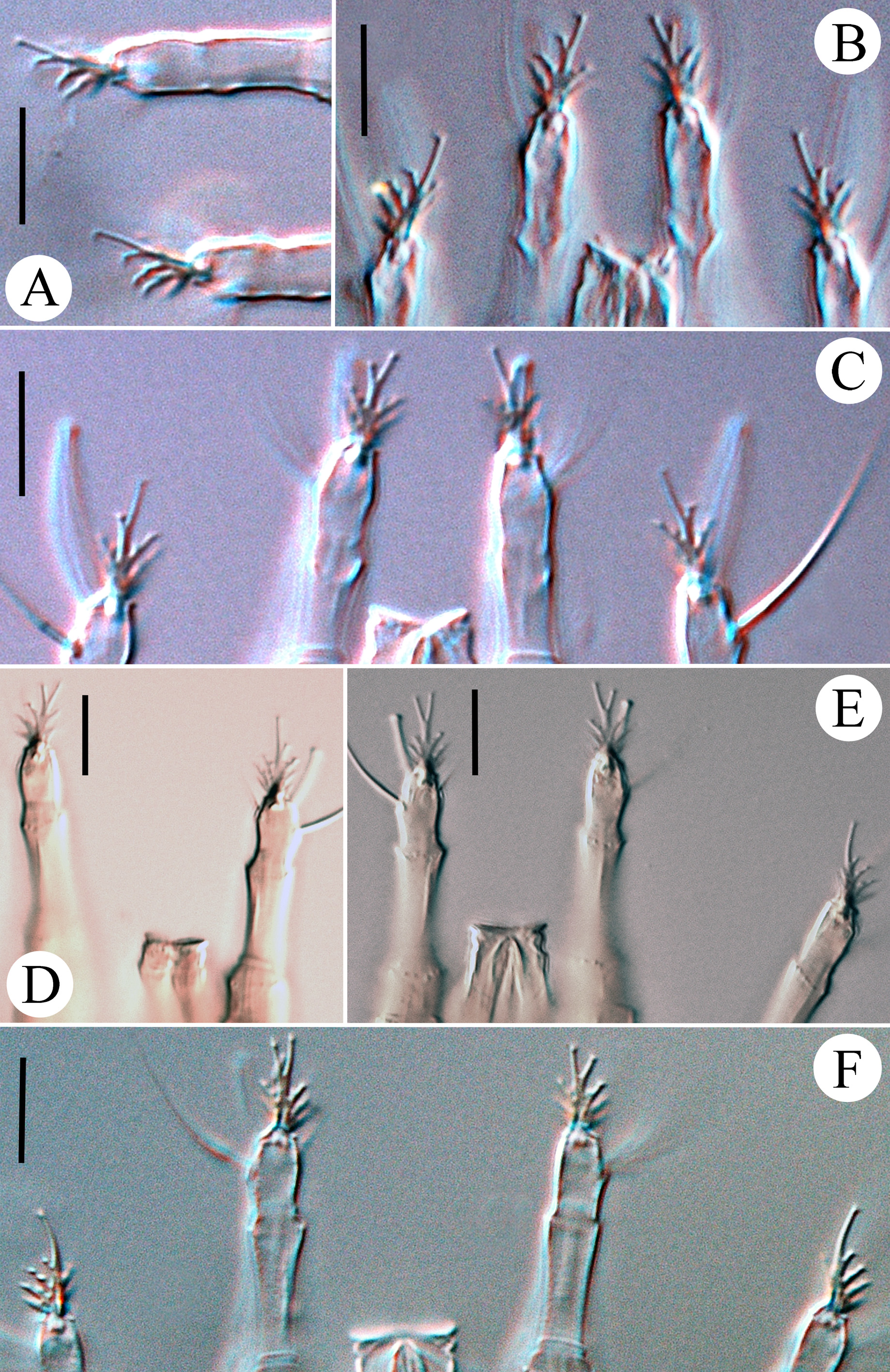
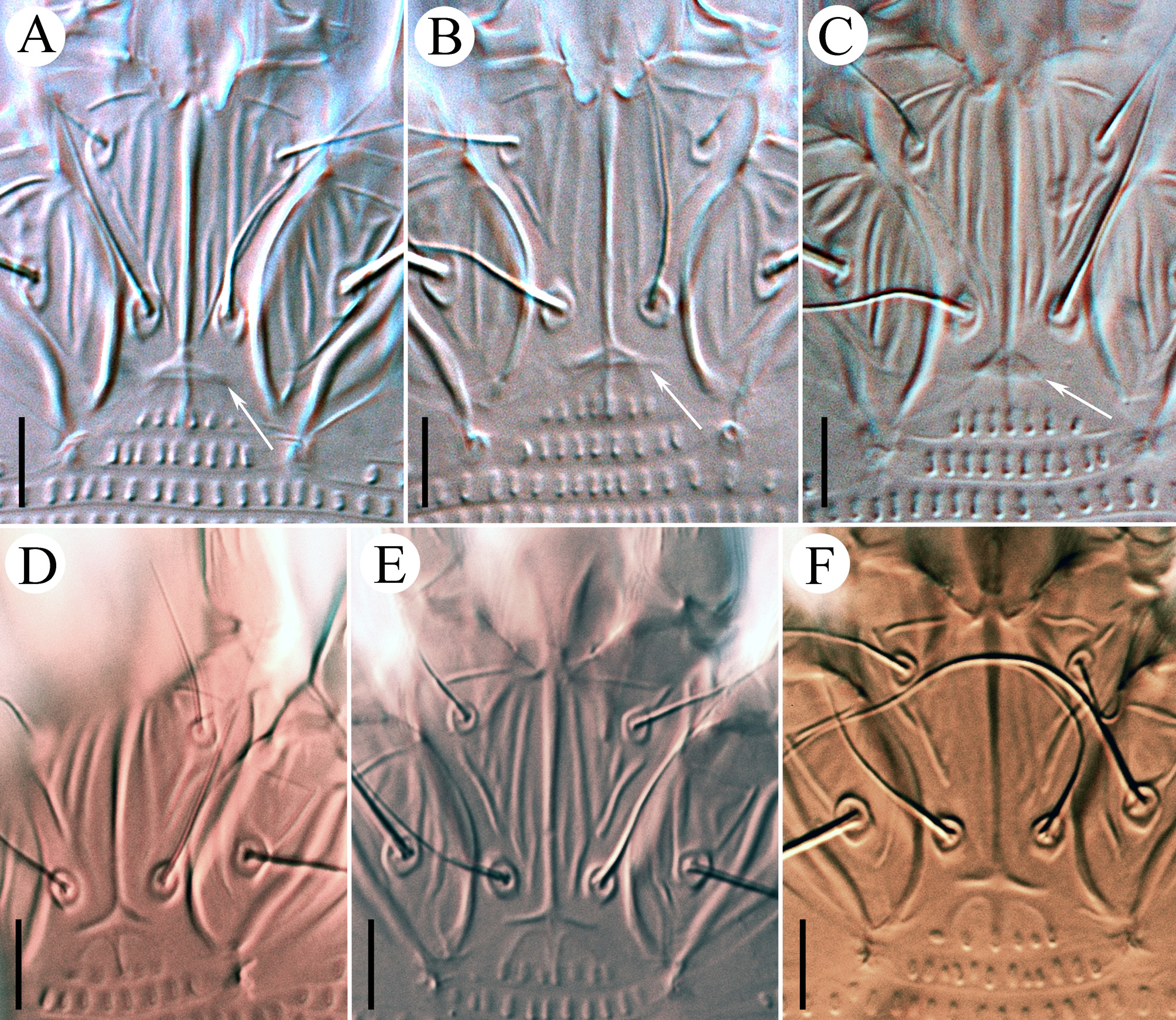
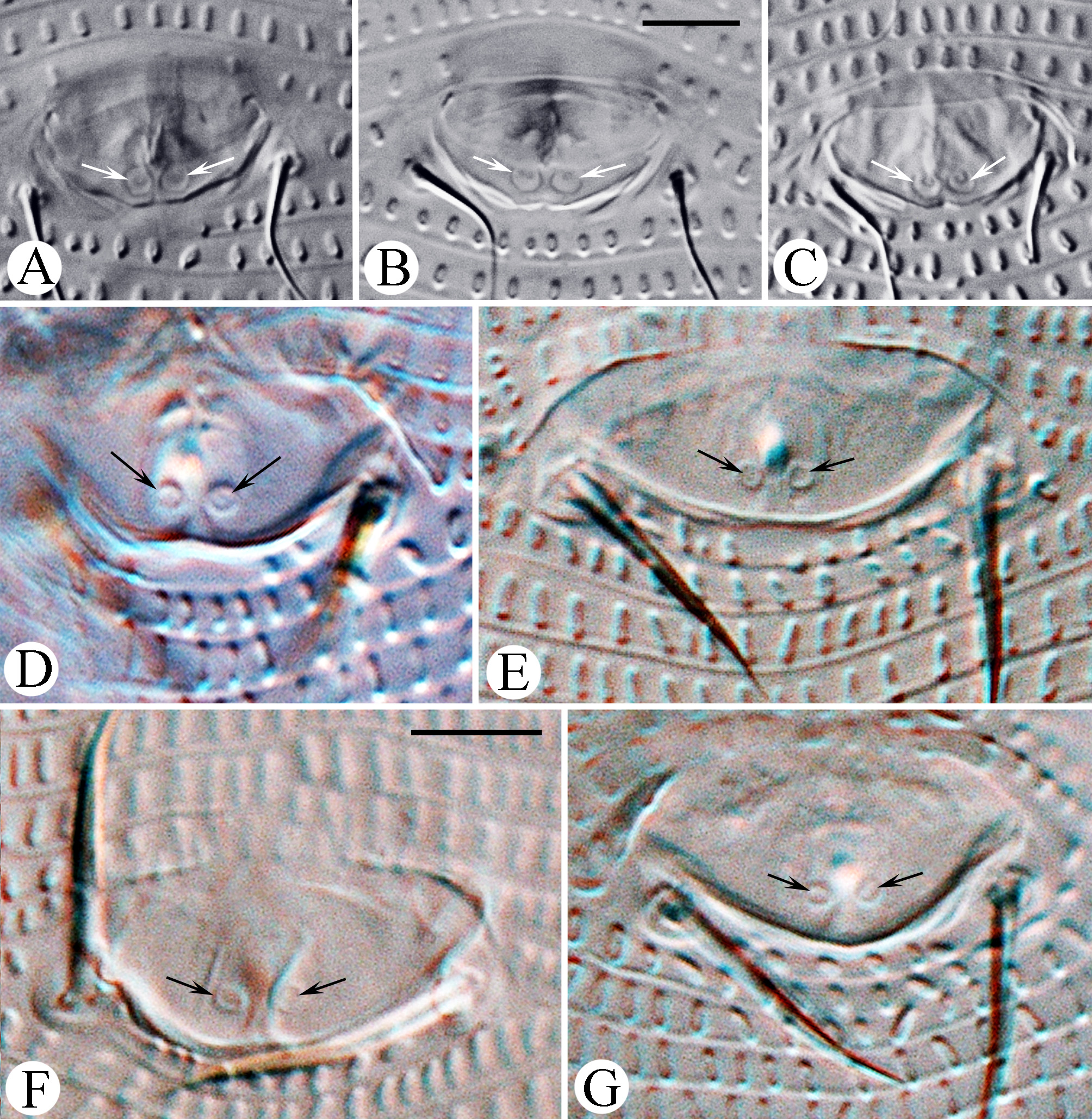
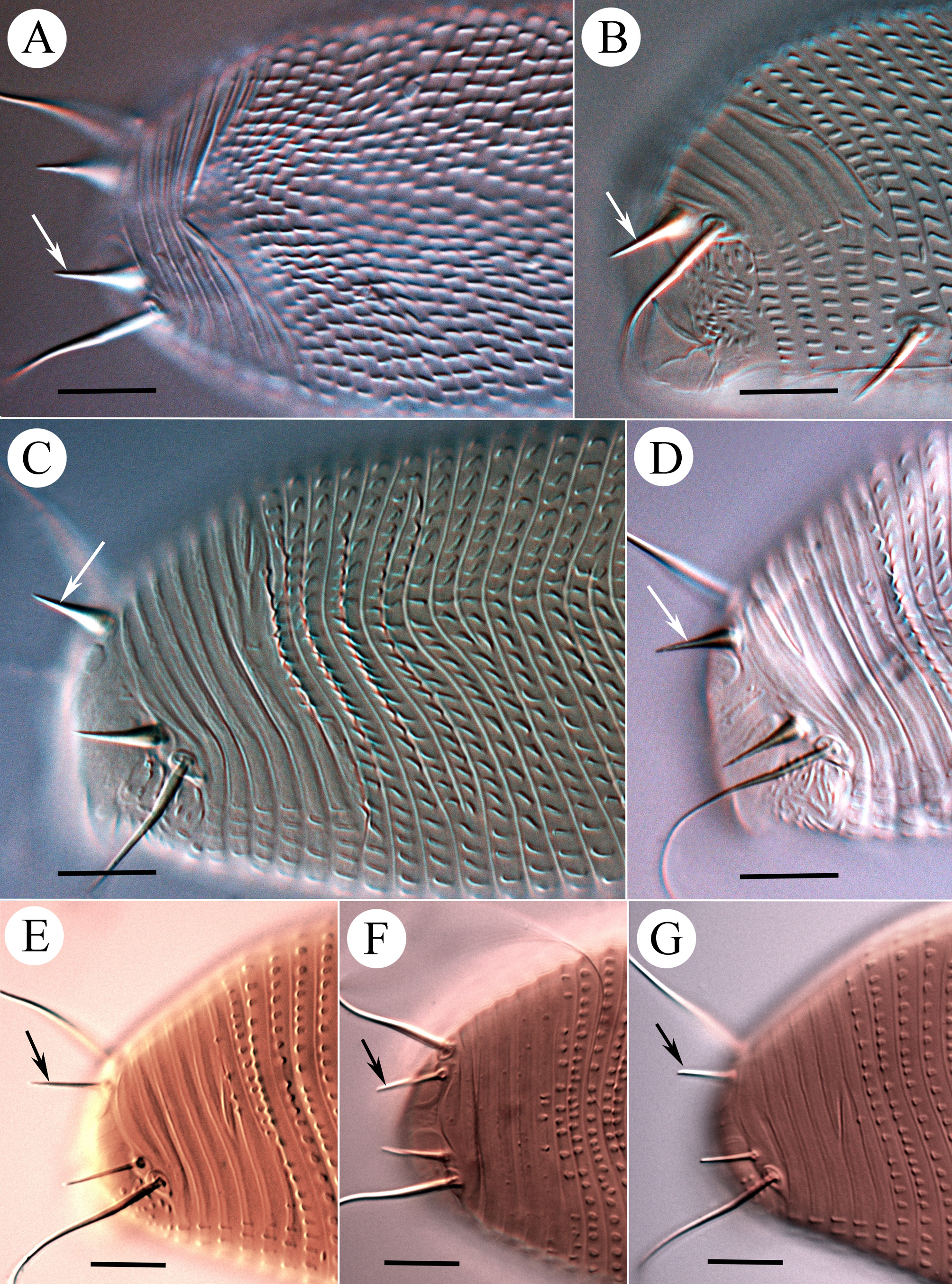
Leg I 38 (32–39), tibia 10 (7–12), l ′ 6 (4–8), tarsus 7 (4–7), u ′ 1 (1–1.5), ft ′ 4 (2–4), ft″ 22 (17–25), ω 5 (4–6); empodium I 4 /4-rayed, 6 (6–8), each ray of the three basal pairs with one additional secondary branch, terminal paired rays without additional branching and usually of equal length (one of the terminal rays of left empodium I was half as long as the other in one specimen; Fig. 2 View FIGURE 2 D). Leg II 9 (6–9), tibia 9 (6–9), tarsus 7 (5–7), u ′ 1 (1–1.5), ft ′ 4 (2–4), ft″ 22 (17–25), ω 8 (7–9); empodium II 4 /4-rayed, 6 (6–8), each ray of the three basal pairs with one additional secondary branch, terminal paired rays without additional branching, internal terminal ray 2–3 times longer than external (thus empodia II are asymmetrical). Tiny denticles present both ventrally and dorsally on anterior margin of trochanter, femur, genu and tibia in legs I and II. Femora I and II with 2–3 ventral longitudinal striae. Coxae each ornamented with 4–5 longitudinal lines. Prosternal apodeme inversely T-shaped, caudally with three short internal processes forming together a tridentate fork ( Fig. 3 View FIGURE 3 D–F). Setae 1b 11 (11–15), 12 (11–15) apart; 1a 17 (14–19), 20 (19–23) apart; 2a 53 (44–54), 20 (19–23) apart; 15 (10–15) coxigenital annuli before epigynium (posterior annuli complete, anterior 2–3 incomplete, i.e. segregated from lateral annuli). Genital coverflap ( Fig. 4 View FIGURE 4 G) semi-circular, smooth, 10 (9–11) x 16 (14–16) wide; setae 3a 13 (11–15), 17 (14–18) apart ( Fig. 6 View FIGURE 6 A–C).
Opisthosoma vermiform, slightly expanded caudally, widest at level of tubercles f; dorsal and ventral annuli bearing small, oval microtubercles, except last 5–6 dorsal annuli devoid of microtubercles. Setal lengths: c2 40 (34–47), d 26 (23–29), e 16 (13–18), f 17 (16–18), h1 11 (8–11), awl-like ( Fig. 7 View FIGURE 7 E–G); h2 broken in type female, (70–105) in paratypes. Setae c2, 3a, d and f, notably thickened. 10 (8–12) annuli from rear shield margin to c2; 14 (12–15) annuli between c2–d; 17 (14–18) annuli between d and e; 25 (20–26) annuli between e and f; 6 (5–6) annuli between f and h1.
Remarks. One aberrant female of N. luzulis n. sp. had a constriction in the central part of the body and an irregular pattern of opisthosomal annuli (slide #u1/3-12) (Fig. 8). This constriction could be considered the result of an unsuccessful nymphal molt, mutation or even perhaps a predator bite-mark. The fact that this female also had deformed tarsi I with shortened, displaced tarsal appendages (not figured here) supports the first two hypotheses.
Male (n=5). Males are shorter than females and have shorter setae sc. They have similarly shaped empodia I and II with terminal rays the same length as basal rays ( Table 1 View TABLE 1 , Fig. 5 View FIGURE 5 I)
Host plant. Luzula pilosa (L.) Willd (hairy wood-rush), distributed in Europe and Siberia.
Relation to the host. Mites live under the leaf epidermis in spaces of mesophyll where they feed on parenchymatous cells. Such feeding putatively causes necrosis of tissues, resulting in a light brown colour of the infested leaf area ( Fig. 9 View FIGURE 9 ).
Type locality. RUSSIA: Arkhangelsk Prov., Plesetsk, 62°43′51″N, 40°17′5″E.
Type material. Female holotype on slide #24-13. In addition, 17 paratype females, six males and ten immatures (each on a separate slide). All specimens from RUSSIA: Arkhangelsk Prov., Plesetsk, 62°43′51″N, 40°17′5″E, 20 August 2012, coll. P.E. Chetverikov.
Additional material. 18 mite specimens on five slide mounts collected 20 August 2004 (same host, locality and collector as type material); 22 mites on 13 slides collected from RUSSIA: Leningrad Prov., Gatchina district, village Belogorka, right bank of river Oredezh, 59°20′32″N, 30°10′29″E, 4 May 2013 and 16 August 2013 (same host and collector).
Etymology. The specific epithet, luzulis , is an adjective, gender masculine, corresponding to the generic name of the host plant.
Differential diagnosis. Mites of N. luzulis n. sp. are morphologically very similar to N. aculeatus Pye, 2012 . These two species have almost identical prodorsal shield ornamentation but differ in the number of dorsal opisthosomal annuli, the shape of h1 and empodia I and II, and lengths of opisthosomal setae c2, d, e, f, which are approximately twice as long in N. luzulis n. sp. as in N. aculeatus ( Table 3 View TABLE 3 ). Additionally, these two species inhabit plants from different juncaceous genera ( Luzula and Juncus , repectively), and their hosts have overlapping distributions: Luzula pilosa is distributed in Europe and Siberia whereas Juncus squarrosus L. (mosquito rush) is native to Europe and Morocco as well as being introduced in the USA (Wisconsin) and Denmark ( Govaerts 2014; USDA NRCS 2014).
Characters Mite species

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.