
Echinoderes leduci, Grzelak & Sørensen, 2022
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.844.1949 |
|
publication LSID |
lsid:zoobank.org:pub:193EDD91-B24D-455C-B8AA-8133586A00A1 |
|
DOI |
https://doi.org/10.5281/zenodo.7225521 |
|
persistent identifier |
https://treatment.plazi.org/id/B0BC5E3C-0F52-4854-B0BD-D80712B6EC30 |
|
taxon LSID |
lsid:zoobank.org:act:B0BC5E3C-0F52-4854-B0BD-D80712B6EC30 |
|
treatment provided by |
Felipe |
|
scientific name |
Echinoderes leduci |
| status |
sp. nov. |
Echinoderes leduci View in CoL sp. nov.
urn:lsid:zoobank.org:act:B0BC5E3C-0F52-4854-B0BD-D80712B6EC30
Figs 11–13 View Fig View Fig View Fig ; Tables 8–9
Diagnosis
Echinoderes with spines in middorsal position on segments 4, 6 and 8 and spines in lateroventral positions on segments 6 to 9. Tubes present in laterodorsal and ventrolateral positions on segment 2, lateroventral positions on segment 5, sublateral positions on segment 8, and laterodorsal positions on segment 9. Sexually dimorphic tubes furthermore present in laterodorsal positions on segment 10 in males. Minute scales present on segments 2 to 10, but regular cuticular hairs absent throughout trunk.
Etymology
The species is named after Dr Daniel Leduc in recognition of his contribution in collecting Hikurangi Margin kinorhynchs and making them available for description to the authors of the present study.
Material examined
Holotype
NEW ZEALAND • ♂; Pahaua Canyon , stn TAN1004/31; 41.4962° S, 175.6828° E; 730 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; NIWA-159405 . Mounted for LM in Fluoromount G on HS slide.
GoogleMapsParatype GoogleMaps
NEW ZEALAND • 1 ♀; Honeycomb Canyon GoogleMaps , stn TAN1004/58; 41.4080° S, 175.8977° E; 670 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; NIWA-159406 . Mounted as holotype.
Additional material
NEW ZEALAND • 1 ♂; Pahaua Canyon , stn TAN1004/27; 41.4983° S, 175.7043° E; 1013 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; personal reference collection of MVS. Mounted for SEM GoogleMaps • 1 ♂; same collection data as for paratype; personal reference collection of MVS. Mounted for SEM GoogleMaps .
Description
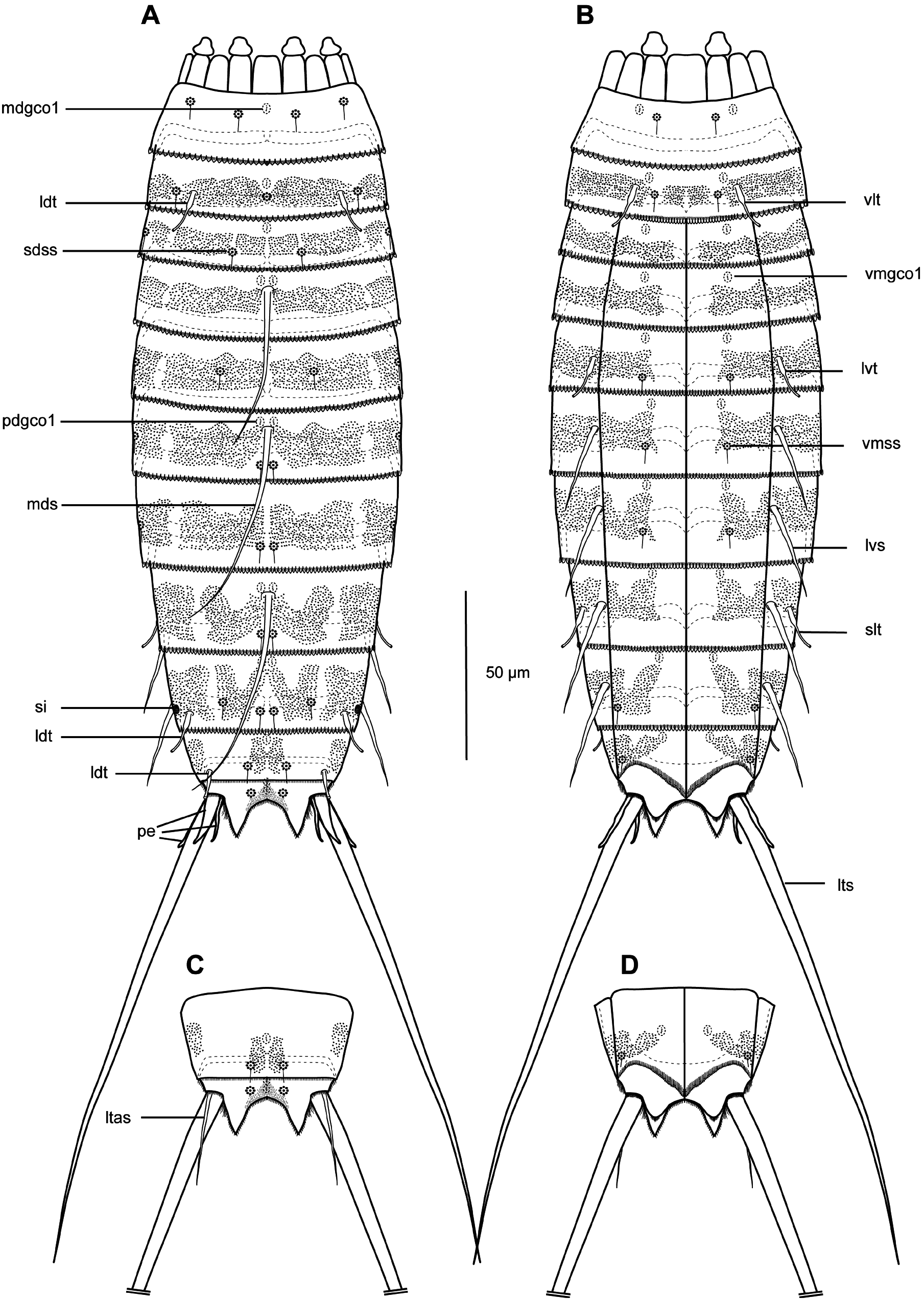
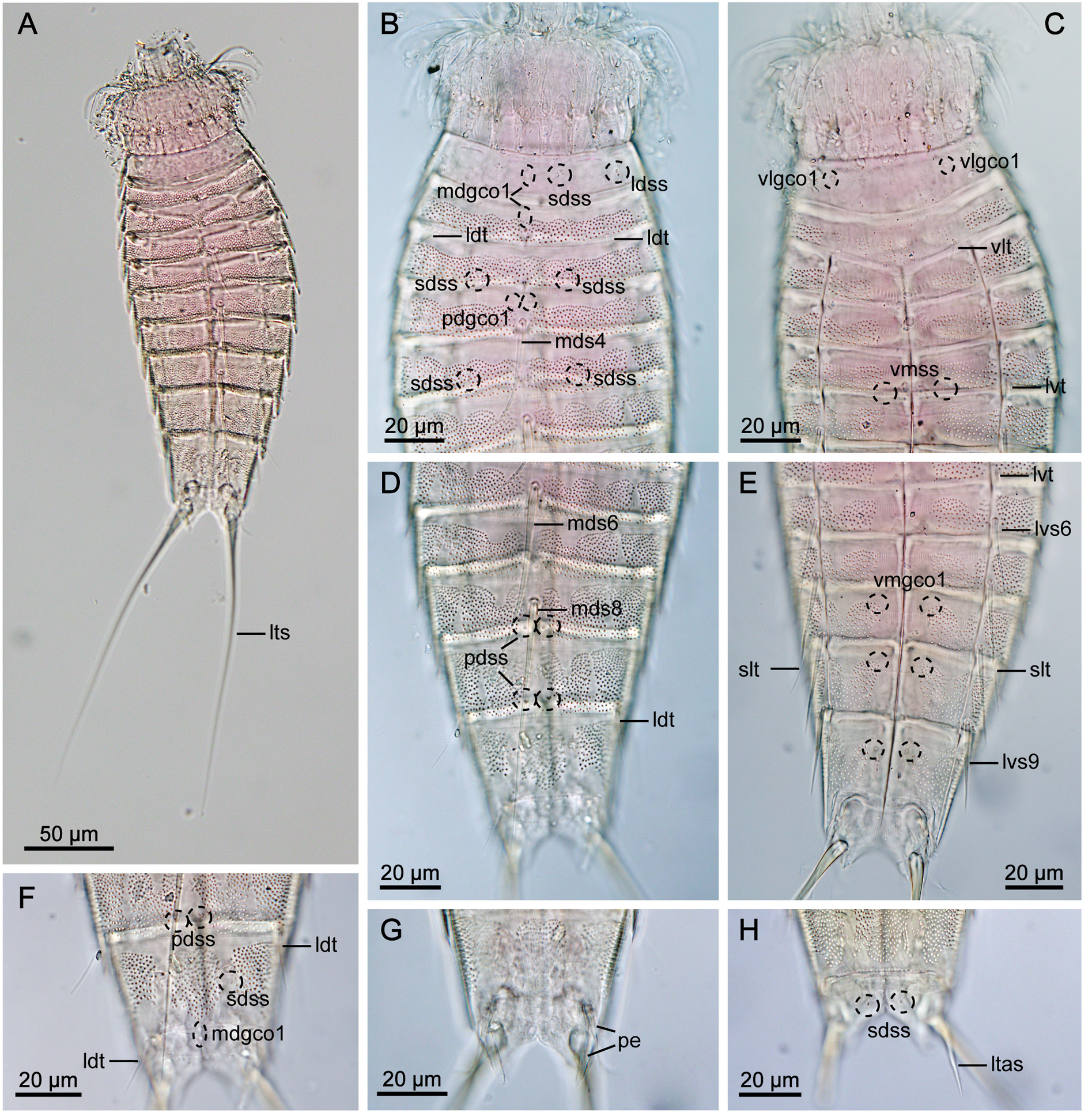
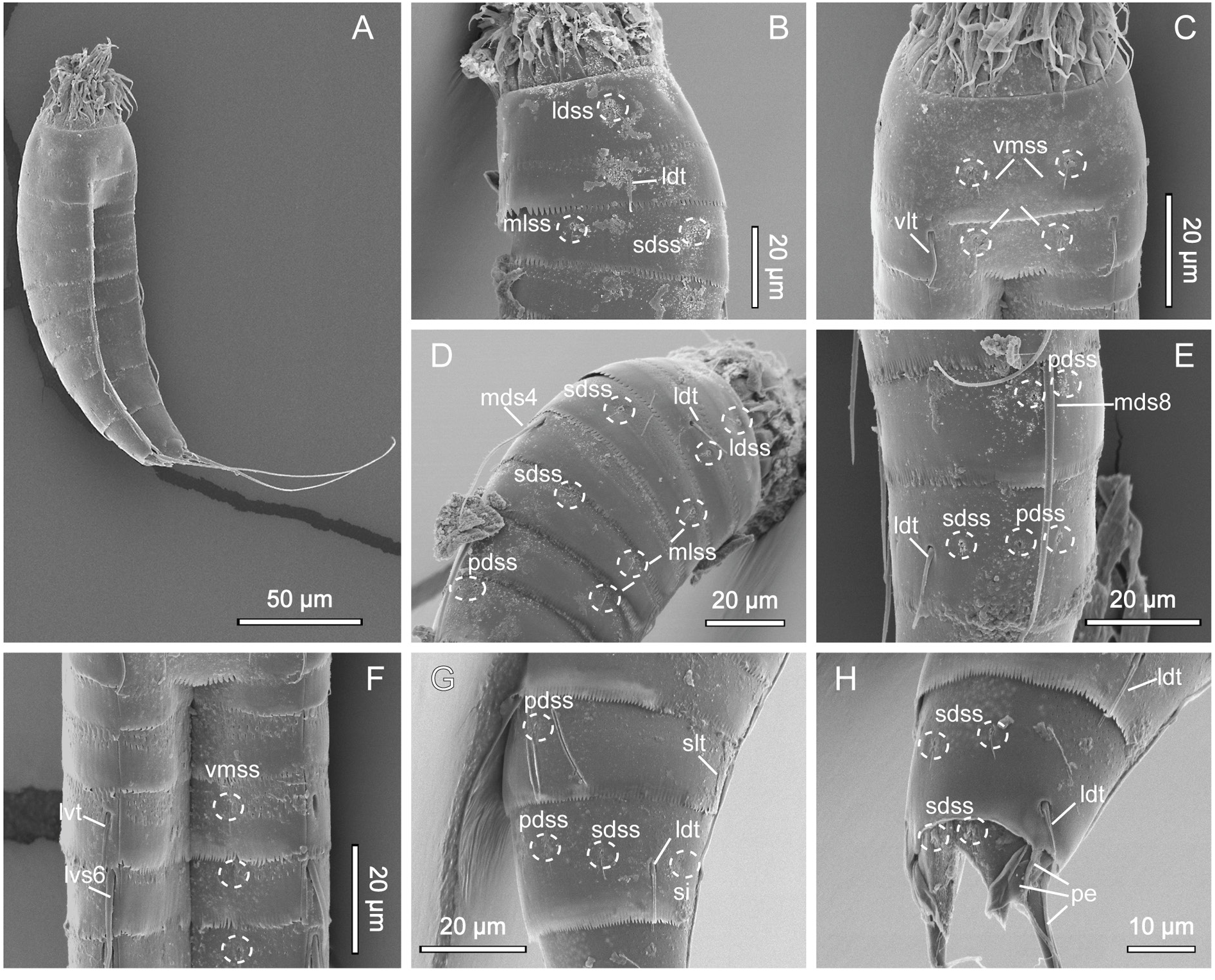
GENERAL. Adults with head, neck and eleven trunk segments ( Figs 11–13 View Fig View Fig View Fig ). Overview of measurements and dimensions in Table 8. Distribution of cuticular structures, i.e., sensory spots, glandular cell outlets, spines and tubes, summarized in Table 9. Head morphology could not be examined in detail in any of available specimens.
NECK. Consists of 16 placids. Midventral placid broadest, 12 µm in width and 14 µm in length, whereas all others narrower, measuring 7 µm in width at their bases ( Fig. 11 View Fig ). Trichoscalid plates well developed.
SEGMENT 1. Consists of complete cuticular ring. Sensory spots present in subdorsal, laterodorsal and ventromedial positions. Sensory spots relatively large with marginal hairs, located on anterior half of segment ( Figs 11A–B View Fig , 12B–C View Fig , 13B–C View Fig ). Glandular cell outlet type 1 present in middorsal position and in ventrolateral positions. Cuticular hairs or perforation sites not present. Posterior segment margin almost straight, forming pectinate fringe with short, sawtooth-like fringe tips ( Fig. 13B–C View Fig ).
SEGMENT 2. Consists of complete cuticular ring, with tubes located in laterodorsal and ventrolateral positions ( Figs 11A–B View Fig , 12B–C View Fig , 13B–C View Fig ). Sensory spots of similar sizes as on preceding segment with long marginal hair, present in middorsal, laterodorsal and ventromedial positions. Glandular cell outlet type 1 present in middorsal position and in ventromedial positions. Pachycyclus of anterior segment margin of regular thickness, interrupted in middorsal position. Secondary pectinate fringe present near anterior segment margin of this and following segments, but mostly covered by preceding segment. This and following nine segments completely hairless. Cuticular hairs reduced to minute scales distributed around segment ( Fig. 13D–G View Fig ), emerging through perforation sites; perforation sites easily visible in LM ( Fig. 12A–H View Fig ). Posterior segment margin almost straight ( Fig. 13C View Fig ); pectinate fringe tips as on preceding segment, except midventral area with slightly narrower fringe tips.
SEGMENT 3. Present segment, and eight remaining ones, consist of one tergal and two sternal plates ( Figs 11A–B View Fig , 12C, E View Fig ). Sensory spots present in subdorsal and midlateral positions. Sensory spots on this and following segments slightly smaller than on preceding segments, but still with one long marginal hair. Glandular cell outlets type 1 present in middorsal and in ventromedial positions. Perforation sites appear as band around segment, interrupted in middorsal and laterodorsal areas and in central part of sternal plate on this and following five segments ( Figs 11A–B View Fig , 12B–E View Fig ). Secondary fringe and posterior segment margin as on preceding segment.
SEGMENT 4. With spine in middorsal position ( Figs 11A View Fig , 12B View Fig , 13D View Fig ). Glandular cell outlets type 1 present in paradorsal and ventromedial positions. No other traits observed. Posterior segment margin straight, terminating in pectinate fringe with slightly slenderer fringe tips along ventral margin than on preceding segments, otherwise as on preceding segment.
SEGMENT 5. With tubes in lateroventral positions ( Figs 11B View Fig , 12C View Fig , 13F View Fig ). Sensory spots present in subdorsal, midlateral and ventromedial positions ( Figs 11A–B View Fig , 12B–C View Fig , 13D, F View Fig ). Glandular cell outlets type 1 present in middorsal and ventromedial positions. Perforation sites, secondary fringe and posterior segment margin as on preceding segment.
SEGMENT 6. With spines in middorsal and lateroventral positions ( Fig. 11A–B View Fig ). Sensory spots present in paradorsal, midlateral and ventromedial positions ( Figs 11A–B View Fig , 13D, F View Fig ). Glandular cell outlets type 1 present in paradorsal and ventromedial positions. Segment otherwise as segment 5.
SEGMENT 7. With spines in lateroventral positions, and sensory spots in paradorsal, midlateral and ventromedial positions ( Figs 11A–B View Fig , 13F View Fig ). Glandular cell outlets type 1 present in ventromedial positions only; no glands observed on dorsal side. Tips of pectinate fringe of posterior segment margin more slender than on preceding segments. Segment otherwise as segment 6.
SEGMENT 8. With spines in middorsal and lateroventral positions, and relatively long tubes (21 µm) in sublateral positions ( Figs 11A–B View Fig , 12D–E View Fig , 13E, G View Fig ). Sensory spots present in paradorsal positions only.
Glandular cell outlets type 1 present in paradorsal and ventromedial positions. Segment otherwise as segment 7.
SEGMENT 9. With spines in lateroventral positions. Long tubes (20 µm) present in laterodorsal positions ( Figs 11A View Fig , 12D, F View Fig , 13E, G View Fig ). Sensory spots located in paradorsal, subdorsal and ventrolateral positions ( Figs 11A–B View Fig , 12E–F View Fig , 13E, G View Fig ). Glandular cell outlets type 1 present paradorsally and ventromedially. Small sieve plates located in midlateral positions ( Fig. 13G View Fig ). Band of perforation site patches interrupted in subdorsal area instead of laterodorsally as on preceding segments. Secondary fringe and posterior segment margin as on preceding segment.
SEGMENT 10. With well-developed laterodorsal tubes, present in males only, located near posterior segment margin ( Figs 11A View Fig , 12F View Fig , 13H View Fig ). Females without tubes ( Figs 11C View Fig , 12H View Fig ). Sensory spots present in subdorsal and ventrolateral positions ( Figs 11A–B View Fig , 12E–F View Fig , 13H View Fig ); subdorsal pair located rather close to paradorsal area. Glandular cell outlets type 1 present in middorsal and in ventromedial positions. Band of perforation site patches as on preceding segment but with additional patch present in paradorsal area. Posterior segment margin of tergal plate straight, without fringe tips ( Fig. 13H View Fig ); margins of sternal plates concave, reaching the posterior margin of the terminal segment, with short and narrow fringe tips.
SEGMENT 11. With lateral terminal spines ( Figs 11A–B View Fig , 12A View Fig , 13A View Fig ). Females with lateral terminal accessory spines ( Figs 11C–D View Fig , 12H View Fig ); males with three penile spines ( Figs 12G View Fig , 13H View Fig ). Dorsal and ventral spines slender and tubular, with ventral ones longer than dorsal ones; median spines very stout, coneshaped ( Figs 11A View Fig , 12G View Fig , 13H View Fig ). Sensory spots present in subdorsal positions. Two unpaired glandular cell outlets type 1 present middorsally. Segment devoid of characteristic perforation sites patches, but with very short cuticular hair-like structures covering paradorsal area. Short fringes covering margins of tergal and sternal plates. Tergal extensions triangular ( Figs 11A, C View Fig , 13H View Fig ). Sternal extensions rounded, not extending beyond tergal extensions ( Fig. 11B, D View Fig ).
Distribution
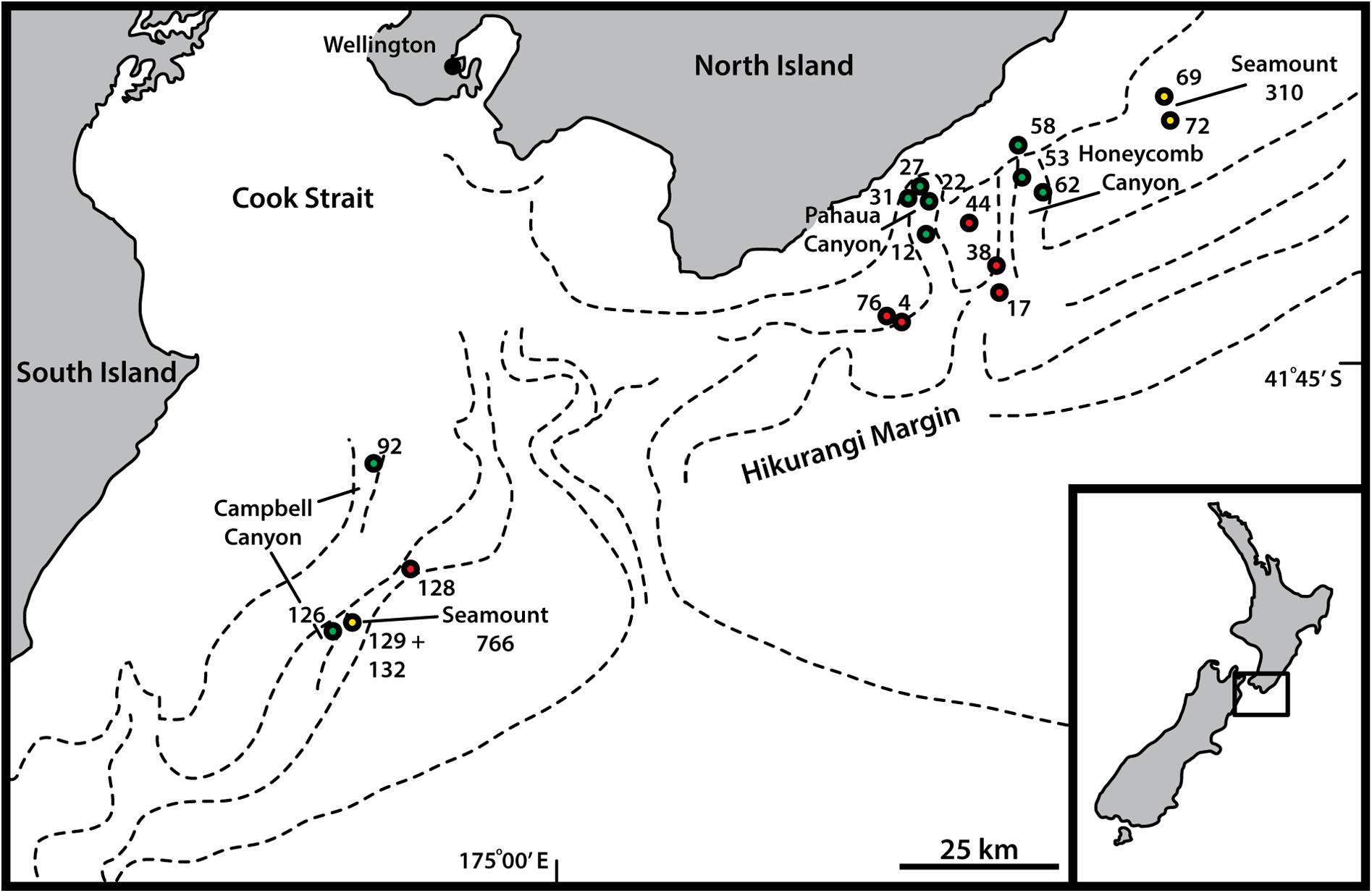
Canyons: Pahaua, Honeycomb, 670–1013 m b.s.l. See Fig. 1 View Fig for a geographic overview of stations and Table 1 View Table 1 for station and specimen information.
Taxonomic remarks on Echinoderes dalzottoi sp. nov. and E. leduci sp. nov.
Echinoderes dalzottoi sp. nov. and E. leduci sp. nov. can easily be recognized by the nature of their trunk appearance with cuticular hairs that are reduced to minute scales, and characteristic strong perforation sites present on segments 2 to 10. The complete lack of cuticular hairs is an uncommon feature among species of Echinoderes . Within Echinoderidae , the species’ peculiar lack of cuticular hairs is shared only with Meristoderes glaber Sørensen et al., 2013 ( Sørensen et al. 2013). Resemblance with M. glaber is only superficial though, and the lack of lateral spines on segment6, together with the presence of glandular cell outlets type 2 instead of tubes on segment 8 in M. glaber easily distinguish it from E. dalzottoi and E. leduci . The new species can also be distinguished from each other without any difficulty. Despite the number of similarities suggesting a close relationship, such as presence of lateroventral spines on segments 6–9, lateroventral tubes on segment 5, sublateral tubes on segment 8 and laterodorsal tubes on segment 9, it is possible to distinguish the two species based on several characters. The main difference is the number of middorsal spines, present on segments 4, 6 and 8 in E. leduci in contrast to spines on segments 4 and 6 only in E. dalzottoi . Furthermore, E. leduci lacks subdorsal tubes on segment 2 (present in E. dalzottoi ), but has laterodorsal tubes on this segment instead.
Having tubes on segment 2 in only laterodorsal and ventrolateral positions as in E. leduci sp. nov. is quite a rare character, and only shared with two other species of Echinoderes , i.e., E. daenerysae and E. higginsi Huys & Coomans, 1989 (although a laterodorsal pair was not mentioned in the original description of the latter species) ( Huys & Coomans 1989; Grzelak & Sørensen 2018). Neither of them can be confused with E. leduci , however. Except for the dense cuticular hair covering observed in both species, E. daenerysae possesses only two middorsal spines on segments 6 and 8, while E. higginsi , although having the same number and arrangement of middorsal spines as E. leduci , is characterized by long, conspicuous spinous tergal extensions of segment 11 and the lack of tubes on segment 9.
Echinoderes dalzottoi sp. nov. is also easily distinguished from all other congeners by its combination of spines and tubes. Middorsal spines being restricted to segments 4 and 6 only is rare and shared only with four other species, i.e., E. astridae , E. bispinosus , E. uozumii , and E. landersi sp. nov. ( Higgins 1982; Sørensen 2014; Yamasaki et al. 2020). However, E. astridae , E. bispinosus and E. uozumii lack dorsal and lateral tubes on segment 2, and have instead two pairs of glandular cell outlet type 2 – a gland type that is absent in E. dalzottoi . Echinoderes landersi , in turn, is characterized by four pairs of tubes on segment 2 and three pairs of tubes on segment 8 – a so far unique tube combination (see ‘Remarks’ in the description of E. landersi above for more details).
The combination of tubes in subdorsal, sublateral and ventrolateral positions on segment 2 is also quite rare. Numerous species present combinations of two or four pairs of tubes on this segment, but only four species have three pairs, among which only E. hispanicus Pardos et al., 1998 and E. peterseni Higgins & Kristensen, 1988 have tube arrangements as in E. dalzottoi sp. nov. ( Higgins & Kristensen 1988; Pardos et al. 1998; Grzelak & Sørensen 2018). However, both species also have three middorsal spines and lack laterodorsal tubes on segment 9, which easily distinguish them from E. dalzottoi . Even considering that sublateral tubes might be absent in some specimens of E. dalzottoi (a variation in the occurrence of tubes has recently been reported for a number of echinoderid species; see, e.g., Grzelak & Sørensen (2018) and Yamasaki & Dal Zotto (2019) for further details), none of the species having only subdorsal and ventrolateral tubes on segment 2 can be confused with E. dalzottoi due to their significantly different middorsal spine patterns and trunk cuticle appearance.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |