
Echinoderes galadrielae, Grzelak & Sørensen, 2022
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.844.1949 |
|
publication LSID |
lsid:zoobank.org:pub:193EDD91-B24D-455C-B8AA-8133586A00A1 |
|
DOI |
https://doi.org/10.5281/zenodo.7225407 |
|
persistent identifier |
https://treatment.plazi.org/id/CD592202-3712-43B6-9F4E-DE5670D87EE7 |
|
taxon LSID |
lsid:zoobank.org:act:CD592202-3712-43B6-9F4E-DE5670D87EE7 |
|
treatment provided by |
Felipe |
|
scientific name |
Echinoderes galadrielae |
| status |
sp. nov. |
Echinoderes galadrielae View in CoL sp. nov.
urn:lsid:zoobank.org:act:CD592202-3712-43B6-9F4E-DE5670D87EE7
Figs 29–31 View Fig View Fig View Fig ; Tables 20–21
Diagnosis
Echinoderes with spines in middorsal position on segments 4 to 8, and spines in lateroventral positions on segments 6 to 9. Tubes present in lateral accessory positions on segment 5. Glandular cell outlets type 2 in subdorsal, laterodorsal, sublateral and ventrolateral positions on segment 2, midlateral positions on segment 5 and sublateral positions on segment 8. Males with minute laterodorsal tubes on segment 10; females with nearly reduced tubes. Segment 11 composed of two tergal and two sternal plates.
Etymology
The species name refers to Galadriel – ‘Lady’ of Lothlórien, one of the greatest of the elves in Middle-Earth, and a character in J.R.R. Tolkien’s “ The Lord of the Rings ” and “ Silmarillion ”. Galadriel helped the Fellowship of the Ring significantly in achieving their goals, hosting them after their escape from the mines of Moria and giving each member a valuable gift for their onward journey.
Material examined
Holotype
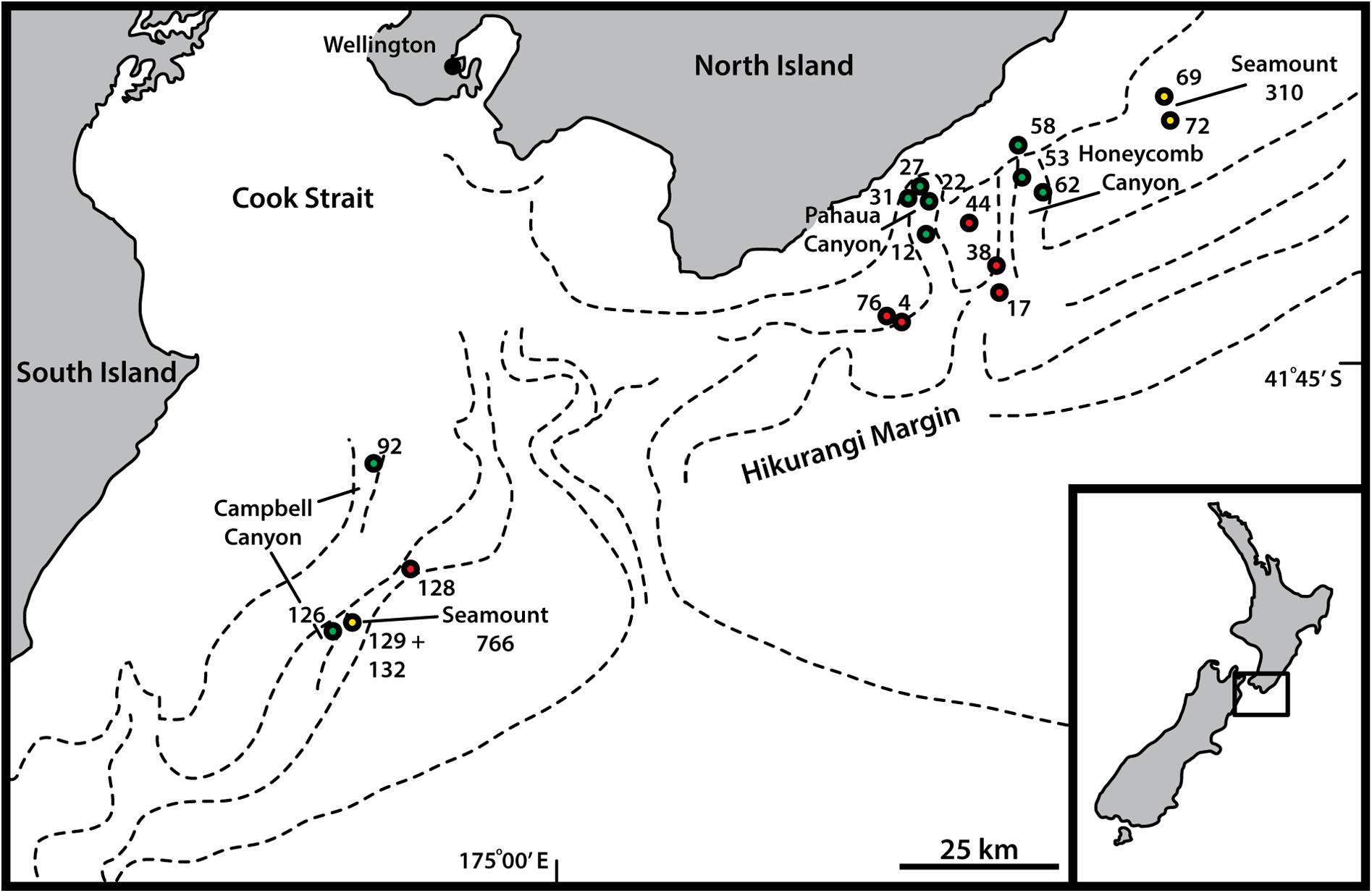
NEW ZEALAND • ♀; Campbell Canyon , stn TAN1004/126; 42.1422° S, 174.5492° E; 1495 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; NIWA-159425 . Mounted for LM in Fluoromount G on HS slide.
GoogleMapsParatypes GoogleMaps
NEW ZEALAND • 2 ♀♀; Pahaua Canyon GoogleMaps , stn TAN1004/12; 41.5508° S, 175.7250° E; 1350 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; NHMD-921496 to 921497. Mounted for LM in Fluoromount G on glass slide • 1 ♀; Seamount 766 , stn TAN1004/132; 42.1345° S, 174.5850° E; 1453 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; NIWA-159426 . Mounted as holotype GoogleMaps .
Additional material
NEW ZEALAND • 2 ♀♀; Hikurangi Slope , stn TAN1004/76; 41.6833° S, 175.6500° E; 1282 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; personal reference collection of MVS. Mounted for SEM GoogleMaps • 3 ♀♀, 1 ♂; Pahaua Canyon , stn TAN1004/27; 41.4983° S, 175.7043° E; 1013 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; personal reference collection of MVS. Mounted for SEM GoogleMaps • 1 ♀; Honeycomb Canyon , stn TAN1004/62; 41.4760° S, 175.9477° E; 1171 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; personal reference collection of MVS. Mounted for SEM GoogleMaps .
Description
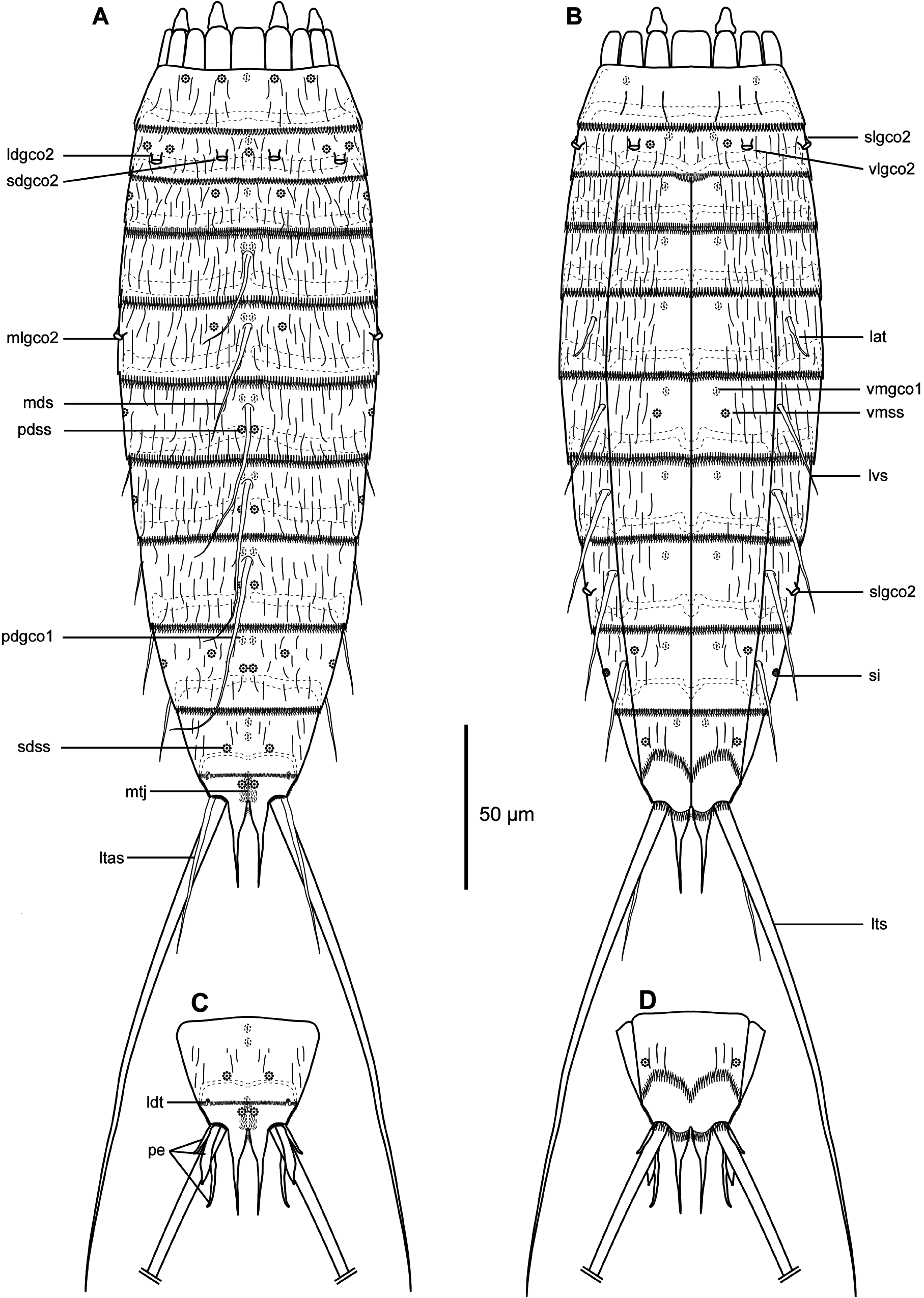
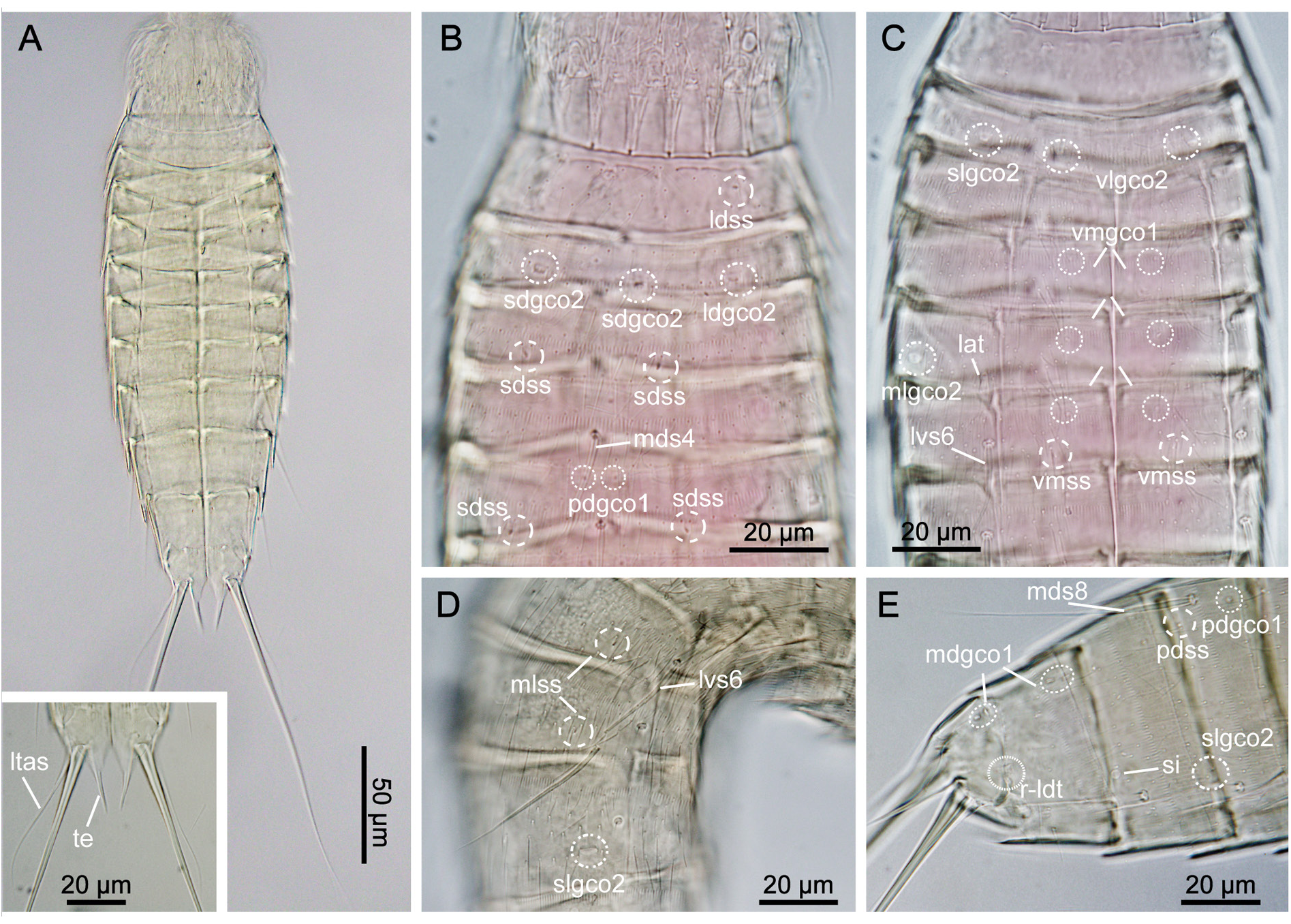
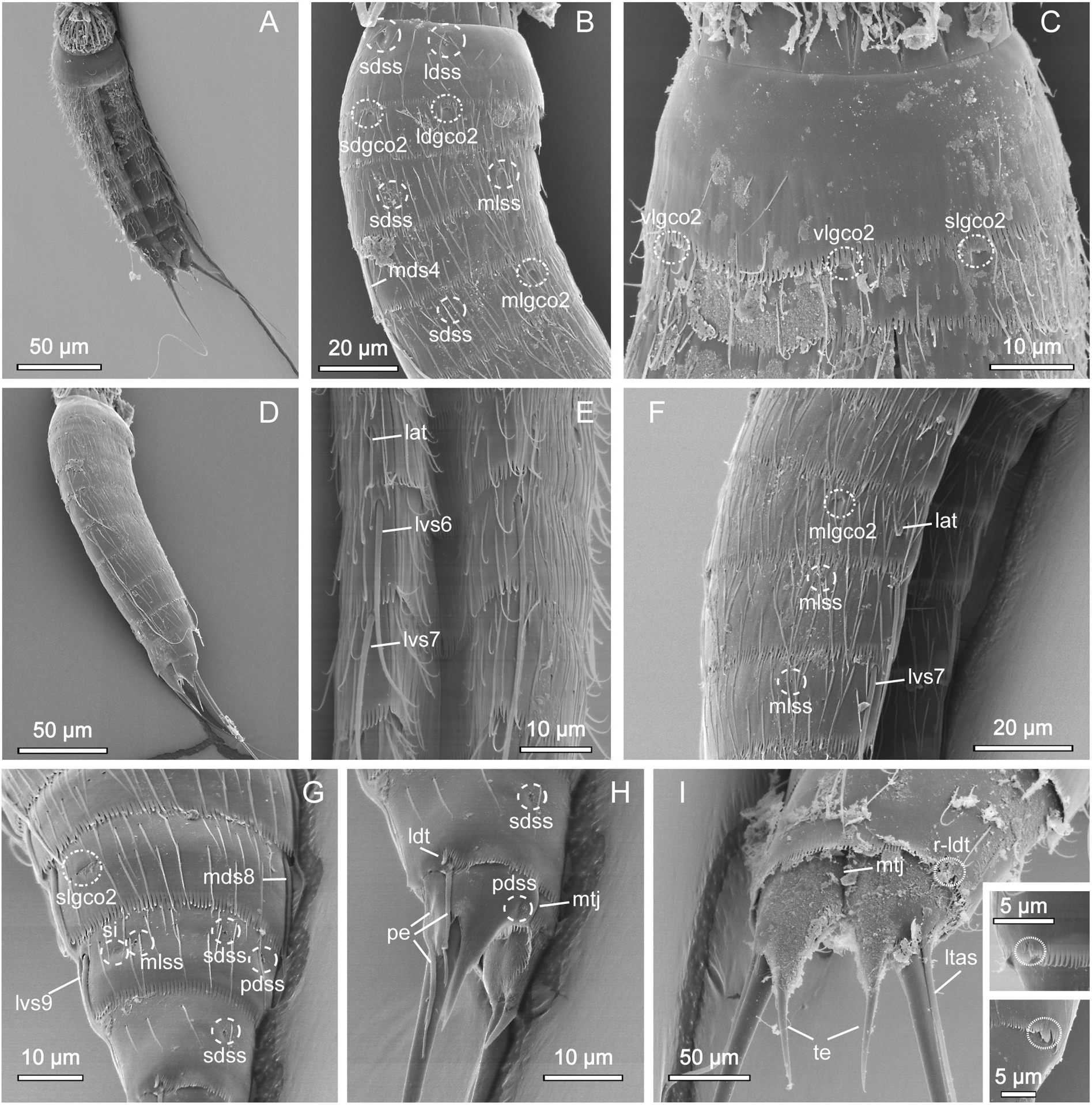
GENERAL. Adults with head, neck and eleven trunk segments ( Figs 29–31 View Fig View Fig View Fig ). Overview of measurements and dimensions in Table 20. Distribution of cuticular structures, i.e., sensory spots, glandular cell outlets, spines and tubes, summarized in Table 21. No details regarding scalid arrangement and morphology could be provided, because introvert of specimens mounted for SEM was fully or partially retracted.
NECK. With 16 placids. Midventral placid broadest, 9 µm in width and 11 µm in length, whereas all others narrower, measuring 6 µm in width at their bases and 11 µm in length, similar in size ( Fig. 30C View Fig , 31D View Fig ). Trichoscalid plates are well developed.
SEGMENT 1. Consists of complete cuticular ring. Sensory spots located anteriorly on segment, in subdorsal and laterodorsal positions ( Figs 29A View Fig , 30B View Fig , 31B View Fig ). Glandular cell outlets type 1 present in middorsal, and ventrolateral positions. Cuticular hairs lightly scattered on dorsal and lateral sides and absent on ventral side ( Fig. 30B–C View Fig ). Posterior segment margin almost straight along dorsal edge, but slightly extended posteriorly in midventral position ( Fig. 31C View Fig ). Pectinate fringe with well-developed fringe tips, slightly shorter and rounded along dorsal margin, and longer and more pointed along lateral and ventral margins ( Fig. 31B–C View Fig ).
SEGMENT 2. Consists of complete cuticular ring. Glandular cell outlets type 2 present in subdorsal, laterodorsal, sublateral and ventrolateral positions ( Figs 29A–B View Fig , 30B–C View Fig , 31B–C View Fig ); glands on ventral side located closer to anterior margin of segment ( Fig. 31C View Fig ). Sensory spots located in middorsal, laterodorsal (two pairs) and ventromedial positions ( Fig. 29A–B View Fig ); sensory spots on this and following segments very small. Glandular cell outlets type 1 present in middorsal and ventromedial positions. Pachycyclus of anterior segment margin of regular thickness, interrupted in middorsal position. Secondary pectinate fringe present near anterior segment margin of this and following segments. Long cuticular hairs lightly scattered on dorsal and lateral sides; ventral side with few hairs. Paraventral and midventral areas in this and following eight segments hairless. Posterior segment margin straight along dorsal side, while extended posteriorly in midventral position. Fringe tips shorter than on preceding segment.
SEGMENT 3. Present segment, and eight remaining ones, consist of one tergal and two sternal plates ( Figs 29A–B View Fig , 30A, C View Fig , 31A View Fig ). Segment with sensory spots located in subdorsal and midlateral positions ( Figs 30B View Fig , 31B View Fig ), and glandular cell outlets type 1 in middorsal and ventromedial positions. Pachycyclus of anterior segment margin of regular thickness, interrupted at tergosternal and midsternal junctions and middorsally, on this and following seven segments. Cuticular hairs more densely distributed across tergal plate, but otherwise as on preceding segment. Posterior segment margin straight, terminating in pectinate fringe with pointed fringe tips, longer than as on preceding segments.
SEGMENT 4. With spine in middorsal position ( Figs 29A View Fig , 31D View Fig ). Segment without sensory spots, but with glandular cell outlets type 1 in paradorsal and ventromedial positions. Pachycycli, pectinate fringe and cuticular hairs as on preceding segment.
SEGMENT 5. With spine in middorsal position and tubes in lateral accessory positions ( Figs 29A–B View Fig , 30C View Fig , 31E–F View Fig ). Glandular cell outlets type 2 present in midlateral positions, being similar in size and shape to those on segment 2. Sensory spots located subdorsally, and glandular cell outlets type 1 present in paradorsal and ventromedial positions. Segment otherwise as segment 4.
SEGMENT 6. With spines in middorsal and lateroventral positions ( Figs 29A–B View Fig , 30A,C View Fig , 31A, D, F View Fig ). Sensory spots present in paradorsal, midlateral and ventromedial positions ( Figs 29A–B View Fig , 30C View Fig , 31F View Fig ). Glandular cell outlets type 1 present in paradorsal and ventromedial positions ( Figs 29A–B View Fig , 30C View Fig ). Tips of pectinate fringe of posterior segment margin as on preceding segments. Cuticular hairs and posterior segment margin as on preceding segment.
SEGMENT 7. With spines in middorsal and lateroventral positions ( Figs 29A–B View Fig , 31D–F View Fig ). Sensory spots present in paradorsal and midlateral positions ( Figs 29A–B View Fig , 31F View Fig ), and glandular cell outlets type 1 in paradorsal and ventromedial positions ( Figs 29A–B View Fig , 30E View Fig ). Cuticular hair covering as on preceding segment. Segment otherwise as segment 6.
SEGMENT 8. With spines in middorsal and lateroventral positions ( Figs 29A–B View Fig , 30E View Fig , 31D, G View Fig ). Glandular cell outlets type 2 located in sublateral positions; gland very conspicuous, larger than those on segments 2 and 5 ( Figs 30D–E View Fig , 31G View Fig ). Sensory spots present in paradorsal positions, and glandular cell outlets type 1 in paradorsal and ventromedial positions ( Fig. 30E View Fig ). Pectinate fringe of posterior segment margin as on preceding segment.
SEGMENT 9. With spines in lateroventral positions ( Figs 29B View Fig , 31A, G View Fig ). Sensory spots located in paradorsal, subdorsal, midlateral and ventrolateral positions; subdorsal pair situated more anterior than others ( Figs 29A–B View Fig , 31G View Fig ). Glandular cell outlets type 1 present in paradorsal and ventromedial positions. Small, rounded sieve plates located in sublateral positions ( Figs 30E View Fig , 31G View Fig ). Pectinate fringe as on preceding segment. Cuticular hair covering and posterior segment margin similar to those on preceding segment.
SEGMENT 10. With sensory spots in subdorsal and ventrolateral positions ( Fig. 31G–H View Fig ). Glandular cell outlets type 1 in ventromedial positions and as pair in middorsal position ( Fig. 30E View Fig ). Males with very short laterodorsal tubes, emerging through slit-like, fringed opening; females with similar slit-like, fringed opening, but with tubes being even further reduced, and either not visible at all or only visible as plate-like projection emerging through opening ( Fig. 31H–I View Fig ). Cuticular hairs very scarce on both tergal and sternal plates ( Fig. 31A, G–I View Fig ). Pectinate fringe of posterior margin on dorsal and lateral sides with markedly shorter fringe tips as on preceding segments, but with longer tips on paraventral areas. Margins of sternal plates extend midventrally.
SEGMENT 11. With pair of lateral terminal spines ( Fig. 29A View Fig ). Females with lateral terminal accessory spines ( Figs 29A View Fig , 30A View Fig , 31I View Fig ); males with three pairs of penile spines ( Figs 29C View Fig , 31H View Fig ), with dorsal- and ventralmost penile spines being slender and tubular, with abrupt narrowings about ¼ from their distal tips; median ones stout and triangular. Sensory spots present in paradorsal positions, near posterior margins of sternal plate ( Fig. 31H View Fig ). A pair of glandular cell outlets type 1 present in middorsal position. Segment devoid of cuticular hairs, but has small patches with short and tiny hair-like extensions in paradorsal positions. Short fringes covering margins of sternal plates. Segment composed of two tergal and two sternal plates ( Fig. 31H–I View Fig ) Tergal extensions significantly elongated and posteriorly projecting ( Figs 30A View Fig , 31H–I View Fig ); sternal extensions short and rounded.
Distribution
Hikurangi Margin, from slope, through canyon, and seamount habitats, 1013–1495 m b.s.l. See Fig. 1 View Fig for a geographic overview of stations and Table 1 View Table 1 for station and specimen information.
Taxonomic remarks on Echinoderes galadrielae sp. nov.
Echinoderes galadrielae sp. nov. displays a very common spine pattern, with five middorsal ones on segments 4 to 8, and lateroventral spines on segments 6 to 9, which is shared with almost 50 species of Echinoderes . However, what makes E. galadrielae differ from most of its congeners is the presence of tubes on segment 5 in lateral accessory positions rather than in lateroventral positions. Such tube displacement is actually a very uncommon trait, observed for only four other species, i.e., E. bathyalis Yamasaki et al., 2018 , E. drogoni Grzelak & Sørensen, 2017 in Grzelak & Sørensen (2018), E. ferrugineus Zelinka, 1928 and E. beringiensis Adrianov & Maiorova, 2022 ( Grzelak & Sørensen 2018; Yamasaki et al. 2018c; Yamasaki & Dal Zotto 2019; Adrianov & Maiorova 2022). Echinoderes galadrielae can easily be distinguished from E. bathyalis though, by its number and positions of glandular cell outlets type 2: E. galadrielae has four pairs of those glands on segment 2, and one pair midlaterally on segment 5 and sublaterally on segment 8, whereas E. bathyalis shows only one pair on segment 2, an absence of glands on segment 5 and lateral accessorily on segment 8 ( Yamasaki et al. 2018c). Moreover, E. bathyalis possesses markedly different tergal extensions, and all its acicular spines are longer.
Echinoderes ferrugineus and E. galadrielae sp. nov. share the same number and arrangement of type 2 glands on segments 2, 5 and 8 (on the latter segment in midlateral rather than sublateral position though), but E. ferrugineus has two additional pairs of glandular cell outlets type 2 on segment 4, in subdorsal and midlateral positions ( Yamasaki & Dal Zotto 2019). Moreover, in addition to its very different tergal extensions, E. ferrugineus also differs by being larger (304 µm vs 224 µm), and having markedly shorter lateroventral and lateral terminal spines (for details see Yamasaki & Dal Zotto 2019).
Among the abovementioned species, two Arctic ones, E. drogoni and E. beringiensis , show the closest resemblance to E. galadrielae sp. nov. The three species share several features, including tube and spine patterns, number and arrangement of glandular cell outlets type 2 and most sensory spots. They also share another very characteristic and uncommon feature, which is the middorsal division of the tergal plate of segment 11 ( Grzelak & Sørensen 2018; Adrianov & Maiorova 2022). Nevertheless, despite overall similarity, E. galadrielae can easily be distinguished from both E. drogoni and E. beringiensis by its elongated tergal extensions.
Such long and conspicuous tergal extensions, constituting almost 15% of the total trunk length, is in fact the most prominent trait of E. galadrielae sp. nov. Longer tergal extensions (TE/TL ~ 20%) are known only for two other species, i.e., E. balerioni Grzelak & Sørensen, 2019 and E. cernunnos Sørensen et al., 2012 (see Sørensen et al. 2012; Grzelak & Sørensen 2019). Echinoderes balerioni can easily be discriminated from E. galadrielae by its three middorsal spines on segments 4, 6 and 8 and the lack of glandular cell outlets type 2. In contrast, E. cernunnos – a species described from Korean waters – shows a much closer resemblance to the new species. The spine and glandular cell outlets type 2 distributions are almost identical with those in E. galadrielae , and a re-examination of the paratypes (NHMD-099881 to 099882) and photos of the holotype (INBRIV-0000245082) of E. cernunnos revealed that both species also have their tubes on segment 5 displaced to lateral accessory positions. The most conspicuous difference between the species is the presence of midlateral glandular cell outlets type 2 on segment 7 in E. cernunnos , which are absent in E. galadrielae . Interestingly, E. cernunnos is also one of the few known congeners (together with abovementioned E. drogoni and E. juliae Sørensen et al., 2018 ) with a middorsal fissure on segment 11 ( Sørensen et al. 2012, 2018; Grzelak & Sørensen 2018) and is also characterised by tergal extensions not only similar in length, but also in their horn-like shape (compare Sørensen et al. 2012: fig. 6f–h with Fig. 31H–I View Fig in the present study). Nonetheless, despite several similarities and the fact that E. galadrielae and E. cernunnos share several rare character traits, telling them apart should not be a problem. Except for the different distribution of sensory spots, the easiest way is to focus on segment 7, which has glandular cell outlets type 2 in E. cernunnos , and on the lateral terminal spines, which are markedly longer in E. galadrielae (LTS/TL=87% in E. galadrielae vs LTS/TL=23% in E. cernunnos ) ( Sørensen et al. 2012).
Hence, in summary, E. galadrielae sp. nov. appears to share a number of traits with E. drogoni , E. cernunnos , and E. beringiensis , including general spine pattern, lateral displacement of the tubes on segment 5, glandular cell outlet type 2 patterns on segments 2, 5 and 8, and middorsal division of the tergal plate of segment 11. This combination of rather unusual characters suggests that the four species are closely related and represent a clade within Echinoderes . Following previous attempts to identify such species groups within the genus (see, e.g., Yamasaki 2016; Sørensen et al. 2018, 2020), we propose that these four species should be considered as a monophyletic entity and referred to as the E. cernunnos species group.
Species with uncertain identities
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |