
Echinoderes aragorni, Grzelak & Sørensen, 2022
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.844.1949 |
|
publication LSID |
lsid:zoobank.org:pub:193EDD91-B24D-455C-B8AA-8133586A00A1 |
|
DOI |
https://doi.org/10.5281/zenodo.7225405 |
|
persistent identifier |
https://treatment.plazi.org/id/88723AE9-DE79-4A1B-840E-3E23F83CA477 |
|
taxon LSID |
lsid:zoobank.org:act:88723AE9-DE79-4A1B-840E-3E23F83CA477 |
|
treatment provided by |
Felipe |
|
scientific name |
Echinoderes aragorni |
| status |
sp. nov. |
Echinoderes aragorni View in CoL sp. nov.
urn:lsid:zoobank.org:act:88723AE9-DE79-4A1B-840E-3E23F83CA477
Figs 26–28 View Fig View Fig View Fig ; Tables 18–19
Diagnosis
Echinoderes with spines in middorsal position on segments 4, 6 and 8, and spines in lateroventral positions on segments 8 and 9. Tubes present in lateroventral positions on segment 5. Glandular cell outlets type 2 present in paradorsal positions on segments 2 and 3, subdorsal and laterodorsal positions on segments 2 to 9, midlateral positions on segments 3 to 5 and 9, sublateral positions on segments 2 and 6 to 8, and in lateral accessory positions on segments 5, 8 and 9. Furthermore, lateroventral glandular cell outlets type 2 present on segments 3, 4, 6, 7, and ventrolateral ones on segment 2. Males with additional pair of glandular cell outlets type 2 in laterodorsal positions on segment 10. A protuberancelike structure emerges between segments 10 and 11 in middorsal position.
Etymology
The species name refers to Aragorn – King of Arnor and Gondor, one of the characters in J.R.R. Tolkien’s “ The Lord of the Rings ”. Aragorn was a confidant of Gandalf, a great warrior and the true leader of the Fellowship of the Ring.
Material examined
Holotype
NEW ZEALAND • ♀; Hikurangi Slope , stn TAN1004/38; 41.5937° S, 175.8532° E; 1121 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; NIWA-159423 . Mounted for LM in Fluoromount G on HS slide.
GoogleMapsParatypes GoogleMaps
NEW ZEALAND • 1 ♀; Hikurangi Slope GoogleMaps , stn TAN1004/4; 41.6837° S, 175.6642° E; 1046 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; NIWA-159424 . Mounted for LM in Fluoromount G on glass slide • 1 ♀; Honeycomb Canyon , stn TAN1004/58; 41.4080° S, 175.8977° E; 1171 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; NHMD-920115 . Mounted as holotype. GoogleMaps
Additional material
NEW ZEALAND • 1 ♀; Hikurangi Slope , stn TAN1004/44; 41.5258° S, 175.8003° E; 728 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; personal reference collection of MVS. Mounted for SEM GoogleMaps • 1 ♂; Honeycomb Canyon , stn TAN1004/62; 41.4760° S, 175.9477° E; 1171 m b.s.l.; Apr. 2010; NIWA TAN1004 Voyage; soft sediment; personal reference collection of MVS. Mounted for SEM GoogleMaps .
Description
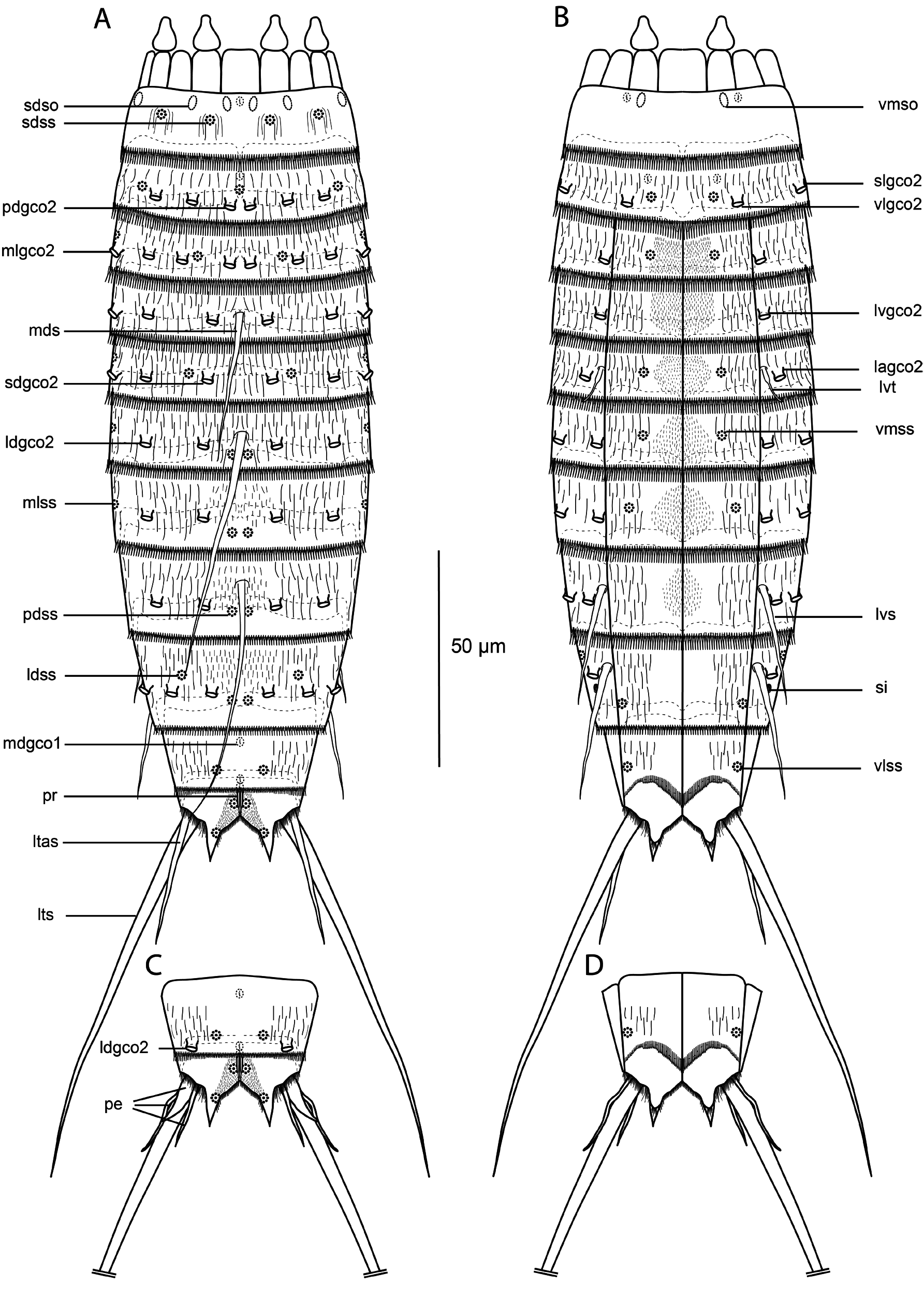
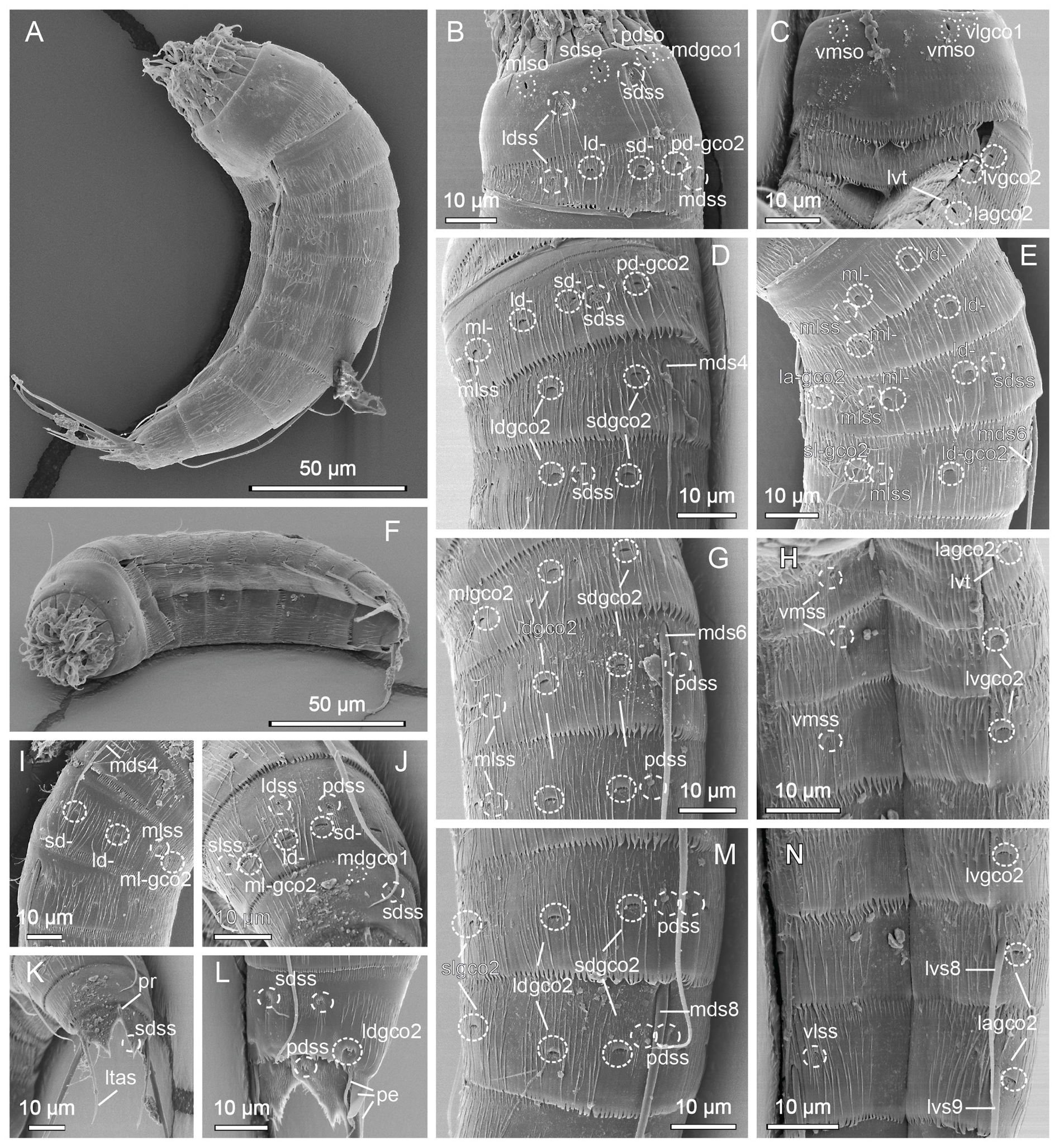
GENERAL. Adults with head, neck and eleven trunk segments ( Figs 26–28 View Fig View Fig View Fig ). Overview of measurements and dimensions in Table 18. Distribution of cuticular structures, i.e., sensory spots, glandular cell outlets, spines and tubes, summarized in Table 19. No details regarding scalid arrangement and morphology could be provided.
NECK. With 16 placids. Midventral placid broadest, 9 µm in width and 10 µm in length, whereas all others narrower, measuring 6 µm in width at bases and 10 µm in length, similar in size ( Figs 27B–C View Fig , 28F View Fig ). Trichoscalid plates well developed.
SEGMENT 1. Consists of complete cuticular ring. Sensory spots located medially on segment, in subdorsal and laterodorsal positions ( Figs 26A View Fig , 27B View Fig , 28B View Fig ). Glandular cell outlets type 1 present in middorsal and ventrolateral positions ( Figs 26A–B View Fig , 27B–C View Fig ). Furthermore, row of unusual cuticular openings present near anterior segment margin in paradorsal, subdorsal, midlateral and ventromedial positions. Openings elongate, or ‘slit-like’, and without any other cuticular characteristic, thus resembling neither sensory spots, flosculi, nor glandular cell outlets of any kind. Due to uncertain nature of these openings, they will simply be referred to as ‘slit-like openings’ ( Figs 26A–B View Fig , 27B–C View Fig , 28B–C View Fig ). Segment devoid of cuticular hairs, except for a few very long hairs arranged around sensory spots. Posterior segment margin almost straight, forming pectinate fringe. Fringe with well-developed long and flexible tips, homogenous along segment margin ( Fig. 28B–C View Fig ).
SEGMENT 2. Consists of complete cuticular ring. Glandular cell outlets type 2 present in paradorsal, subdorsal, laterodorsal, sublateral and ventrolateral positions ( Figs 26A–B View Fig , 27B–C View Fig , 28B, D View Fig ). Sensory spots in middorsal, laterodorsal and ventromedial positions ( Figs 26A–B View Fig , 27B View Fig , 28B, D View Fig ). Glandular cell outlets type 1 present in middorsal and ventromedial positions. Pachycyclus of anterior segment margin of regular thickness, without interruption. Secondary pectinate fringe present near anterior segment margin of this and following segments. Cuticular hairs evenly distributed across segment. Posterior segment margin almost straight; pectinate fringe tips as on preceding segment.
SEGMENT 3. Present segment, and eight remaining ones, consist of one tergal and two sternal plates ( Figs 26A–B View Fig , 27C, E–F View Fig , 28D View Fig ). Pachycyclus of anterior segment margin of regular thickness, interrupted at tergosternal and midsternal junctions and middorsally, on this and following seven segments. Segment with glandular cell outlets type 2 in paradorsal, subdorsal, laterodorsal, midlateral and lateroventral positions ( Figs 26A–B View Fig , 27B–C View Fig , 28C–E View Fig ). Sensory spots in subdorsal, midlateral and ventromedial positions ( Figs 27B View Fig , 28D–E View Fig ). Glandular cell outlets type 1 not observed. On this and following three segments cuticular hairs evenly distributed across tergal plate, except for hairless areas located close to midlateral line; ventromedial and paraventral areas on this and following five segments with thinner and much shorter, non-bracteate hairs. Posterior segment margin straight, terminating in pectinate fringe with fringe tips as on preceding segments.
SEGMENT 4. With spine in middorsal position ( Figs 26A View Fig , 27B View Fig ). Glandular cell outlets type 2 present in subdorsal, laterodorsal, midlateral and lateroventral positions ( Figs 26A–B View Fig , 27B–C View Fig , 28D–E View Fig ). Sensory spots or glandular cell outlets type 1 not present. Pachycycli, pectinate fringe and cuticular hairs as on preceding segment.
SEGMENT 5. With tubes in lateroventral positions ( Figs 26B View Fig , 27C View Fig , 28C, H View Fig ). Glandular cell outlets type 2 located in subdorsal, laterodorsal, midlateral and lateral accessory positions ( Figs 26A–B View Fig , 27C, E View Fig , 28E,G–H View Fig ). Sensory spots present in subdorsal, midlateral and ventromedial positions; in holotype and other female specimens midlateral sensory spots located closer to sublateral line, on outer/lateral side of midlateral glandular cell outlets type 2 ( Fig. 28E View Fig ), while in male specimen this pair of sensory spots on dorsal side of midlateral glands ( Fig. 28I View Fig ). Segment otherwise as segment 4.
SEGMENT 6. With spine in middorsal position only ( Figs 26A–B View Fig , 27A, E View Fig , 28A, G View Fig ). Glandular cell outlets type 2 present in subdorsal, laterodorsal, sublateral and lateroventral positions ( Figs 26A–B View Fig , 27C, E View Fig , 28G–H View Fig ). Sensory spots located in paradorsal, midlateral and ventromedial positions ( Figs 26A–B View Fig , 28G– H View Fig ). Tips of pectinate fringe of posterior segment margin as on preceding segments. Segment otherwise as segment 5.
SEGMENT 7. Without spines or tubes ( Figs 26B View Fig , 27E View Fig ). Glandular cell outlets type 2 and sensory spots as on segment 6 ( Figs 26A–B View Fig , 27E–F View Fig , 28G–H View Fig ), but ventromedial pair of sensory spots located closer to ventrolateral line than on preceding segment( Fig. 28H View Fig ). Cuticular hair covering as on segment 6, except for paraventral areas with much shorter and thinner hairs than on preceding segments.
SEGMENT 8. With spines in middorsal and lateroventral positions ( Figs 26A–B View Fig , 27G View Fig , 28E View Fig ). Glandular cell outlets type 2 present in subdorsal, laterodorsal, sublateral and lateral accessory positions ( Figs 26A–B View Fig , 27E–F View Fig , 28M–N View Fig ). Sensory spots present in paradorsal positions only ( Fig. 28M View Fig ). Pectinate fringe of posterior segment margin with slightly narrower and shorter tips on the dorsal side. Segment otherwise as segment 7.
SEGMENT 9. With spines in lateroventral positions ( Figs 26B View Fig , 27F View Fig , 28N View Fig ). Glandular cell outlets type 2 present in subdorsal, laterodorsal, midlateral and lateral accessory positions ( Figs 26A–B View Fig , 27E–F View Fig , 28J, N View Fig ); subdorsal pair situated close to paradorsal area ( Fig. 28J View Fig ). Sensory spots present in paradorsal, laterodorsal, sublateral and ventrolateral positions; laterodorsal pair located more anteriorly on segment than other pairs ( Figs 26A–B View Fig , 28J View Fig ). Small, rounded sieve plates located in lateral accessory positions ( Fig. 27F View Fig ). Pectinate fringe with shorter tips than on preceding segment, homogenous along segment margin. Cuticular hair covering of tergal plate similar to that on preceding segment, but with broader area of short and thin non-bracteate hairs, covering paradorsal and subdorsal areas; sternal plates devoid of hairs in paraventral and ventromedial areas.
SEGMENT 10. With laterodorsal glandular cell outlets type 2 located near posterior segment margin in males; females without glandular cell outlets type 2 on this segment ( Figs 26A View Fig ,,C, 28K–L). Sensory spots present in subdorsal and ventrolateral positions ( Fig. 28L View Fig ). Glandular cell outlet type 1 present in middorsal position. Cuticular hairs sparser than on preceding segment. Central part of tergal plate almost completely hairless, sternal plate hair covering as on preceding segment.Posterior segment margin of tergal plate straight, with much shorter fringe and narrower tips than those on preceding segment ( Fig. 28K– L View Fig ); margins of sternal plates extend midventrally ( Fig. 26B View Fig ).
SEGMENT 11. With pair of lateral terminal spines ( Fig. 27D, F View Fig ). Males with three pairs of penile spines; dorsal and ventral penile spines thin and flexible tubes, whereas median ones markedly thicker, conical and stout ( Figs 26C View Fig , 28L View Fig ); females with lateral terminal accessory spines ( Figs 26A View Fig , 27F View Fig , 28K View Fig ). Sensory spots present in paradorsal and subdorsal positions; subdorsal pair located near posterior margin of tergal extensions ( Figs 26A View Fig , 28K View Fig ). Pair of glandular cell outlets type 1 present in middorsal position, covered by middorsal protuberance-like structure extending from intersegmentary joint ( Figs 26A,C View Fig , 28K–L View Fig ). Segment devoid of cuticular hairs, but with dense covering of short hair-like extensions on dorsal areas of tergal plates. Short fringes covering margins of tergal and sternal plates; fringe tips slightly longer near insertion of lateral terminal spines. Tergal extensions triangular, sternal extensions do not extend beyond tergal extensions.
Distribution
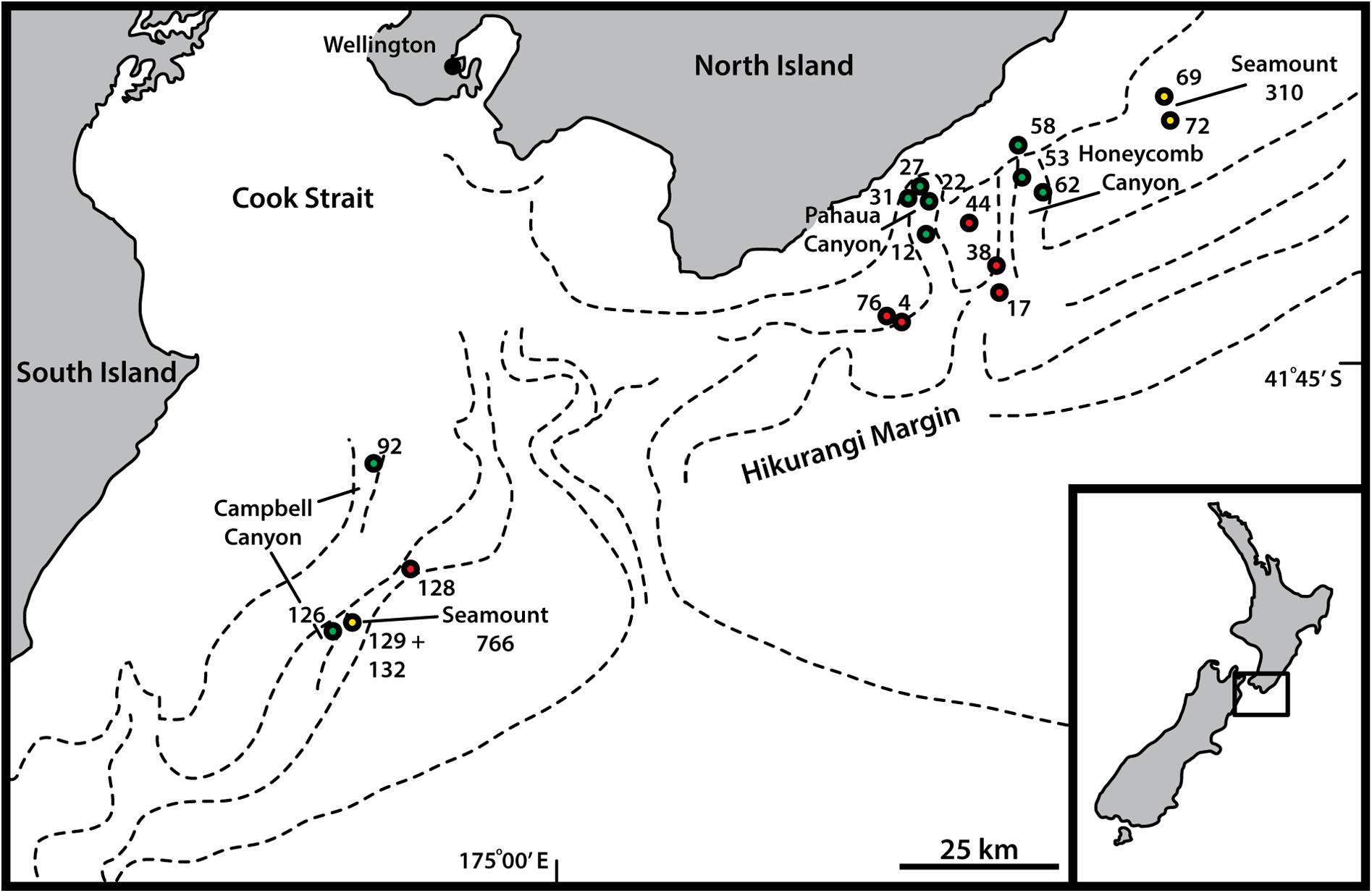
Hikurangi slope and Honeycomb Canyon, 670–1171 m b.s.l. See Fig. 1 View Fig for a geographic overview of stations and Table 1 View Table 1 for station and specimen information.
Taxonomic remarks on Echinoderes aragorni sp. nov.
Echinoderes aragorni sp. nov. can very easily be distinguished from all other congeners by the number and arrangement of glandular cell outlets type 2. No less than 34 pairs of glandular cell outlets type 2 (35 in males) are present in E. aragorni , which is not present in any other kinorhynch species. The only other species displaying a somewhat similar trait is E. orestauri Pardos et al., 2016 with 30 fringed tubules distributed on segments 2 to 9 ( Pardos et al. 2016a). It seems likely that glandular cell outlets type 2 and fringed tubes are homologous, but they still differ morphologically since only the fringed tubes form actual external appendages. In contrast, the glands in E. aragorni are relatively large openings (although smaller in male specimens than in females: see Fig. 28I View Fig vs Fig. 28J View Fig ), easily distinguishable in both SEM and LM and present on segments 2 to 10.
A second rare feature in E. aragorni sp. nov. is the presence of lateroventral tubes/spines on segments 5, 8 and 9 only. This feature is shared with three other species only: E. caribiensis Kirsteuer, 1964 , E. lusitanicus and E. skipperae Sørensen & Landers, 2014 ( Kirsteuer 1964; Neves et al. 2016; Sørensen & Landers 2014). However, none of these species can be confused with E. aragorni , as they do not possess even a single pair of glandular cell outlets type 2. Moreover, E. caribiensis belongs to the E. coulli species group and is characterized by the absence of middorsal spines, while in contrast E. lusitanicus has five middorsal spines. Only E. skipperae has, similar to the new species, a middorsal spine arrangement with spines present on segments 4, 6 and 8.
The presence of slit-like openings on segment 1 is also unique to E. aragorni sp. nov. Other species show an occurrence of cuticular structures on segment 1 like tubes or glandular cell outlets type 2 (e.g., E. frodoi sp. nov., E. legolasi sp. nov., E. anniae , and E. hamiltonorum ) ( Sørensen et al. 2018; present study), but the structures observed in E. aragorni have not been observed before. Although it may represent another type of glandular cell outlets – or a modified kind of glandular cell outlets type 2 – we are currently uncertain about their nature and function. Therefore, it is difficult to discuss this observation further, but it is obviously a structure that deserves more attention in future works.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |