
Brachyscelus crusculum Spence Bate, 1861
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5026.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:11E41F28-9FA8-4057-93E2-59D6538DA631 |
|
persistent identifier |
https://treatment.plazi.org/id/03B487A3-FFA1-4617-F3A0-FAFBFDE877AC |
|
treatment provided by |
Plazi |
|
scientific name |
Brachyscelus crusculum Spence Bate, 1861 |
| status |
|
Brachyscelus crusculum Spence Bate, 1861 View in CoL
( Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Brachyscelus crusculum Spence Bate, 1861: 7–10 View in CoL , pl. 2, figs 1–2.— Spence Bate, 1862: 333–335, pl. 53, figs 2–3.— Claus 1873: 468.— Claus 1876: 519.— Bovallius 1887a: 31.— Stebbing, 1888: 1544–1549, pls. 195–196.— Chevreux 1893: 70–74, text figs 1–9.— Chevreux 1900: 153.— Norman 1900: 134.— Tattersall 1906: 26.— Holmes 1908: 490.— Walker 1909: 54.— Sexton 1911: 223.— Stewart 1913: 262.— Spandl 1924: 32.— Chevreux & Fage 1925: 427–428, fig. 418.— Stephensen 1925: 172–176, 230 (table).— Schellenberg 1927: 649–650, fig. 49.— Spandl 1927: 210.— Pirlot 1929: 139–140.— Barnard 1930: 432.— Pirlot 1930: 25–26.— Barnard 1932: 292.— Boone 1935: 222–226, pl. 66.— Chevreux 1935: 196–197.— Pirlot 1939a: 46–47.— Dakin & Colefax 1940: 123, fig. 212.— Shoemaker 1945: 242.— Irie 1948: 36 (table).— Shoemaker 1948: 13.— Bulycheva 1955: 1048 (table).— Hurley 1955: 181 (key).— Reid 1955: 25–26.— Irie 1957a: 10 (table).— Irie 1957b: 355, fig. 23.— Irie 1959: table 3.—Nayer 1959: 47.— Hurley 1960: 282, 285 (table).— Evans 1961: 200.— Kane 1962: 310– 311.— Vinogradov 1962: 23–24.— Siegfried 1963: 6 (list), 10.— Sheard 1965: 244 (list).— Pillai 1966: 225–226, fig. 15.— Sheard 1967: passim.— Hure et al. 1969: 603 & 605 (tables).— Hurley 1969: 33, pl. 19 (map 8).— Dick 1970: 68–69, fig. 13 (part).— Yoo 1971: 43 (list), 62, fig. 23 (map).— Brusca 1973: 9 (table), 19.— Tashiro & Jossi 1972: fig. 8 (map), 20 (list), 34–35 (table).— Thurston 1976: 388–389 (table), 433–434, 463 & 464 (tables), figs 18B, 24 (distribution).— Harbison et al. 1977: 472.— Madin & Harbison 1977: 457.— Shulenberger 1977: 379 (table).— Tranter 1977: 649 (table), 650.— Zeidler 1978: 28–29, 49, figs 28–29.— Laval 1980: 20 & 23 (tables).— Shulenberger 1980: passim.— Brusca 1981: 31 (key), 44, figs 18g, 18j, 18m. — Vinogradov et al. 1982 /1996: 396/489 (key), 396–398/489–492, fig. 213.— Zeidler 1984: passim.— Nagata 1986: 274–275.— Young & Anderson 1987: 717 (table), 719, 721.— Barkhatov & Vinogradov 1988: passim.— Lin & Chen 1988: 325.— Young 1989: 717 (table).—Vinogradov 1990: 77, 95 (table).— Vinogradov 1991: 261 (table), 264.— Zeidler 1991: 133.— Zeidler 1992: 115–116 (incl. key), fig. 19.— De Broyer & Jażdżewski 1993: 118 (list).— Vinogradov 1993: 45 (table).— Lin & Chen 1994: 115, 118 (list).— Montú 1994: 132 (list).— Lin et al. 1995: 118, 122 (table).— Shih & Chen 1995: 178 (key), 178–180, figs 116–117.— Tarling et al. 1995: 34 (table).— Lin et al. 1996: 231 (table).—Vicencio & Fernández 1996: 23 (list).— Montú 1998: 599.— Zeidler 1998: 96.— Barkhatov et al. 1999: 808 (table).— Vinogradov 1999: 1145 (table), 1193 (incl. key), fig. 4.134.— Lowry 2000: 325 (list).— Gasca & Shih 2001: passim.— Lima & Valentin 2001: 473 (list), 474 (table).— Escobar-Briones et al. 2002: 367 (list).— Gasca 2003a: 308 (table).— Gasca 2003b: 118 & 119 (tables).— Gasca & Shih 2003: 95 (table).—Gasca 2004: 997 & 998 (tables).— Gasca & Haddock 2004: 532 (table), 533, fig. 2f. — Gasca & Suárez-Morales 2004: passim.— Vinogradov et al. 2004: 15–16, 24 (table).— Brusca & Hendrickx 2005: 151 (list).— Browne et al. 2007: 819 (table), fig. 4 (phylogenetic tree).— Gasca 2007: 118 (table).— Potier et al. 2007: 65 & 67 (tables).— Gasca & Franco-Gordo 2008: 569 (table).— Costa et al. 2009: 103, 197–198, fig. 187.— Gasca 2009a: 89 (table), 91.— Gasca 2009b: 66 (table).— Gasca et al. 2009: 1497 & 1499 (tables).— Lavaniegos & Hereu 2009: passim.— LeCroy et al. 2009: 969 (table).— Zeidler & De Broyer 2009: 63–64.— Bucklin et al. 2010: 2239 (table).— Mori et al. 2010: 10 (list).— Gasca & Morales-Ramírez 2012: 229, fig. 3D.—Gasca et al. 2012: passim.—Hurt et al. 2013: 31 (table), figs 1, 2 (phylogenetic).— Nunes et al. 2013: passim.— Valencia et al. 2013: 51 (table).—Gasca & Franco- Gordo 2014: 76 (list).— Zhang et al. 2014: passim.— Burridge et al. 2016: passim, fig. 1 (part).— Souza et al. 2016: 344 (table).— Zeidler 2016: 47 (incl. key), figs 16–17.— Gasca & Browne (2017): 3 (table), 6–7, fig. 2d.— Hereu et al. 2020: passim.— Espinosa-Leal et al. 2021a: passim.— Espinosa-Leal et al. 2021b: passim.— Véliz et al. 2021: passim.
Brachyscelus mediterranea View in CoL — Senna 1903: 5–8, figs 1–10.
Brachyscelus mediterraneus — Chevreux 1900: 154.
Brachyscelus acuticaudatus Stebbing, 1888: 1555–1556 View in CoL , pl. 197C.— Grice & Hart 1962: 300.
Brachyscelus stebbingi Boone, 1935: 226–230 View in CoL , pls. 67, 68.
Type material. Type material of Brachyscelus crusculum could not be found in any major European institution and is considered lost. Spence Bate (1861) refers to an ovigerous female “length ¾ of an inch” (about 19 mm). The type locality is unknown; not recorded.
Type material of synonyms. Thamyris mediterranea Claus, 1887 is here regarded a junior synonym of B. globiceps (see under that species). However, the description and figures provided by Senna (1903) are characteristic of B. crusculum . The record of Chevreux (1900) also most likely refers to this species.
The unique holotype female of Brachyscelus acuticaudatus is in the NHM, London (89.5.15.306), on two microscope slides. No precise type locality is given, just the Pacific, Challenger, August or September 1875. An examination of this holotype has confirmed the synonymy of Stebbing’ species. Indeed, the pointed telson, as illustrated by Stebbing (1888), is characteristic of larger females of B. crusculum .
The unique holotype female of Brachyscelus stebbingi (about 10.5 mm) is in the Vanderbilt Marine Museum. The type locality is the north-east Pacific, the Canary Islands, off Fuerte Ventura, Alva, 1931, 250 fathoms. Pereopods 5 and 6 and the telson, as illustrated by Boone (1935), are characteristic of B. crusculum .
Material examined. Type material of B. acuticaudatus as detailed above and the following. In AM: Tasman Sea, off eastern Australia, 139 females, 112 males (7 lots, P39850–6). In Canadian Museum of Nature: tropical Atlantic, just west of Barbados, 41 females, 33 males (34 lots). In NHM: S.E. and mid-Atlantic, 17 females, 10 males (9 lots, 1936.11.2.3243–57), Discovery collections as detailed by Barnard (1932). S. Pacific, near New Zealand, 2 females, 6 males, several juveniles (12 lots, 1930.8.1.754–65), Terra Nova collections as detailed by Barnard (1930). In NHMD: numerous specimens from the Mediterranean Sea and N.E Atlantic collected by the Thor as detailed by Stephensen (1925). Gulf of Panama, Galathea station 745, one female, 3 males. S. Pacific Kermadec deep, Galathea station 671, one male. Tasman Sea, 2 females, 3 males, 4 juveniles, Galathea stations 551, 552, 574, 601, 607. Tropical Atlantic, 2 females, 4 males, Galathea stations 24, 98, 110. In SAM & SAMA (part): Meiring Naude collections from S.E. Indian Ocean, off South Africa, between Kosi Bay and just south of East London, 508 females, 209 males and numerous juveniles (116 lots), mostly 200–0 m. In SAM: S.W. Atlantic, off South Africa, 22 females, 13 males (11 lots). In SAMA: N.E. Indian Ocean, off northern Western Australia, Ningaloo Reef, 13 females, 6 males (8 lots, C14075–14082). Coral Sea, Great Barrier Reef, 15 females, 4 males, 3 juveniles (14 lots, C14047–14059). Tasman Sea, off eastern Australia, 168 females, 79 males, 11 juveniles (78 lots). Great Australian Bight, off South Australia [35°07’51”S 130°41’42”E], 100–0 m, 5 females (C12169). In USNM: numerous specimens from the N.W. and tropical Atlantic and the tropical Pacific.
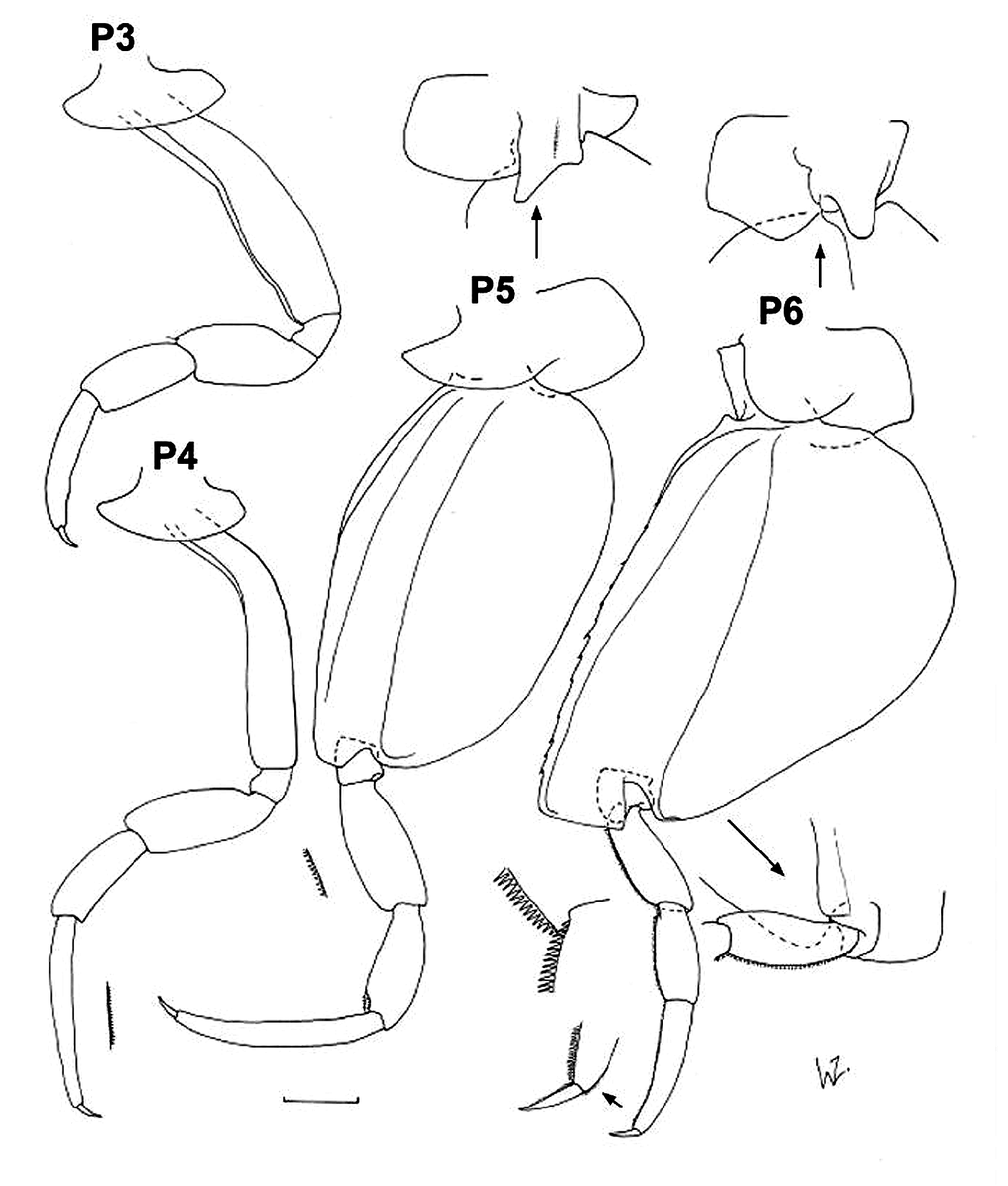
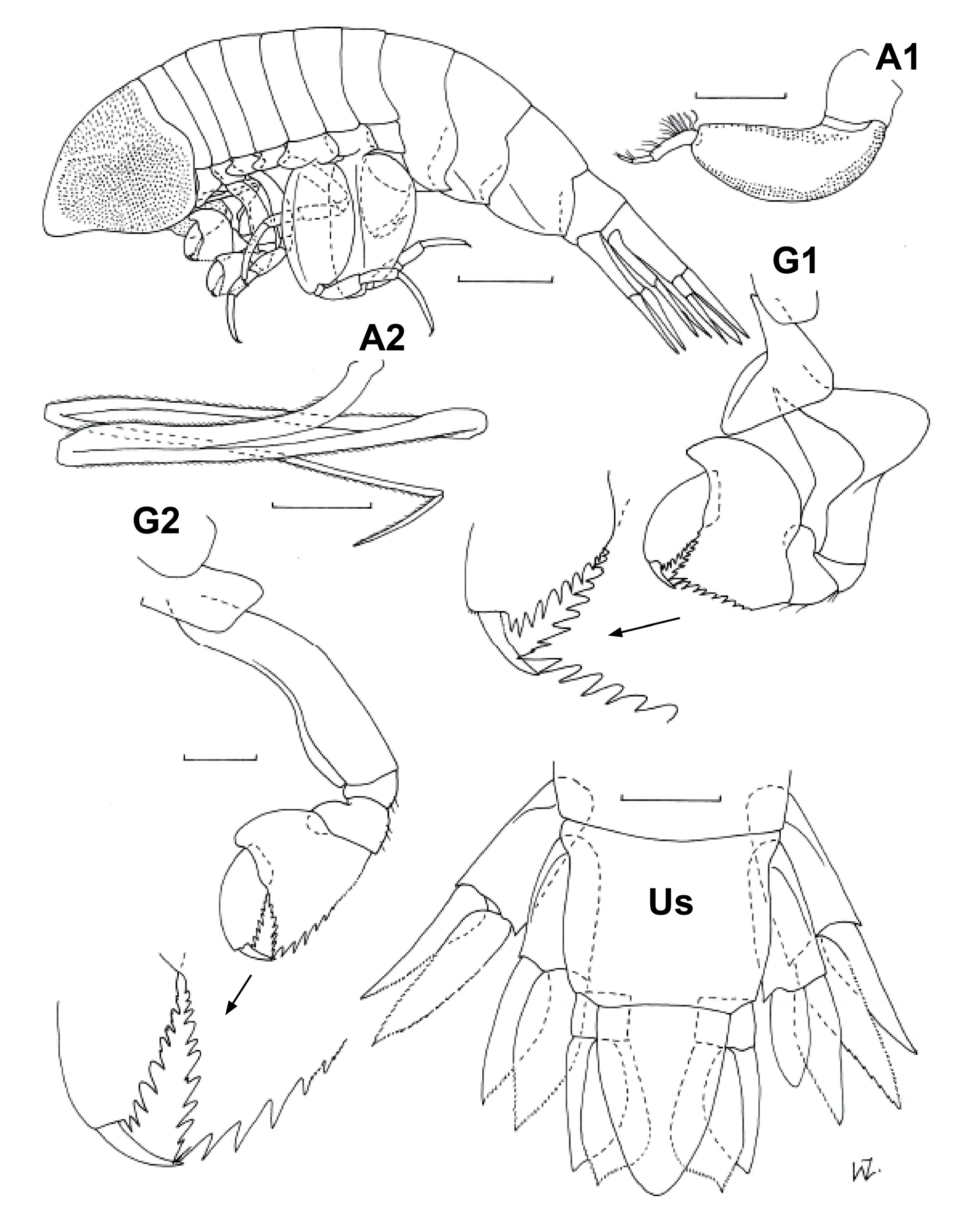
Diagnosis. Body length up to 21 mm for females, 18 mm for males. Head globular in females, depth about 1.5 x length; produced to rounded point in males, marginally longer than deep; more pointed than for other species of the genus. Antennae 2 of males with terminal article shorter than one-third of preceding article. Gnathopod 1 basis slightly shorter and more robust than for G2, with distinct pocket for A 2 in males; carpus with antero-distal corner produced into small lobe, partly overlapping propodus, especially in males; carpal process with large teeth, mainly without very minor serrations; postero-distal corner of propodus armed with one smaller tooth followed by slightly larger teeth of similar size; dactylus relatively sharp, length about 0.4 x propodus. Gnathopod 2 similar to G1 but basis without pocket and antero-distal corner of carpus barely produced. Pereopods 3 and 4 morphologically similar, P4 marginally longer than P3; merus slightly inflated anteriorly, length about 0.4–0.5 x basis; carpus slightly shorter than merus; propodus 1.3 x carpus length for P3 and 1.8 x carpus for P4, posterior margin slightly serrated. Pereopod 5 marginally longer than P6; basis oval-shaped, length almost twice maximum width, antero-distal lobe with relatively straight distal margin, overlapping about half of ischium; merus about 0.3 x basis length, slightly longer than carpus, anterior margin with slight serrations; propodus about 1.8 x carpus length; dactylus relatively short, about 0.2 x propodus. Pereopod 6 basis length 1.5 x maximum width for females, 1.7 x for males, anterior margin straight, antero-distal lobe relatively broad with straight distal margin produced just beyond ischium; posterior margin with distinct bulge proximally tapering gradually to small rounded lobe distally overlapping ischium; merus about 0.3 x basis length (slightly more for males), slightly longer than carpus; propodus about 1.8 x carpus length; anterior margin of merus, carpus and propodus with small serrations; dactylus relatively short, about 0.2 x propodus. Pereopod 7 basis with posterior margin inflated proximally, length about 1.5 x maximum width, almost twice as long as remaining articles combined; merus about 2 x carpus length; propodus about halflength carpus, with complex projections on antero-distal corner; dactylus hook-shaped, partly retractile. Uropod 1 endopod slightly longer than peduncle and exopod. Uropod 2 endopod slightly longer than peduncle, marginally longer than exopod. Uropod 3 endopod about 3.0 x peduncle length, marginally longer than exopod, broadened distally, with distinct excavation terminally on inner margin, slightly broader in males. Rami of all uropods with serrated margins. Double urosomite slightly wider than long. Telson length about 1.5 x width at base, apex pointed in mature females, more rounded in males and immature females.
Remarks. This is the most common and largest species of Brachyscelus and many of the other species have been confused with it in the past, thus some of the above records may be erroneous. Distinctive characters of B. crusculum are: i) males have a pointed head; ii) the shape of the basis of pereopod 6 has a broad, relatively large, antero-distal lobe with a straight distal margin, and a smaller posterior lobe, both of which cover the ischium; iii) the double urosomite is almost as long as it is broad; and iv) the very broad endopod of uropod 3 is especially broad in mature specimens.
In recently moulted males, recognised by the undeveloped antennae, the head is rounded as in females but becomes pointed later ( Zeidler 1992: fig. 19). Also, in freshly moulted males, the antennal pocket of the first gnathopods is not very noticeable but becomes deeper as the second antennae develop and the head becomes more pointed. These morphological changes during moulting and growth may have contributed to past mis-identifications of this and other species of Brachyscelus .
Amongst the material examined (SAM, SAMA) are many smaller specimens, mature at about 7 mm from the Indian Ocean off South Africa, off north Western Australia, the Coral Sea and the Tasman Sea, which may represent a new species, but which could not be distinguished from typical B. crusculum . Males are generally typical of B. crusculum and cannot be distinguished from very large specimens, more than twice the size (fig. 3), collected from the same general area. Females sometimes have a relatively larger head, almost approaching that of B. macrocephalus , but other diagnostic characters match those of B. crusculum except that the telson is not always as acuminate, but this may be a feature of smaller or immature specimens. Stephensen (1925) recorded ovigerous females at 14–20 mm from the Mediterranean Sea but mentions one ovigerous female of the “mediterranean form” at 7 mm. Clearly there is scope for further taxonomic analysis of the “ B. crusculum ” clade (and the genus) using more sophisticated methods such as molecular analysis of fresh specimens from a wide range of geographical regions.
Brachyscelus crusculum has been recorded in association with salps ( Stephensen 1923, 1925; Young &Anderson 1987) and medusae ( Pirlot 1939a), specifically the salps Cyclosalpa affinis ( Chamisso, 1819) , Soestia zonaria ( Pallas, 1774) (previously in Iasis ), Pegea socia ( Bosc, 1802) , Salpa maxima Forsskål, 1775 , Thalia democratica ( Forsskål, 1775) ( Harbison et al. 1977) and Salpa fusiformis Cuvier, 1804 ( Laval 1980) , and the medusa Aequorea coerulescens (Brandt, 1835) ( Gasca & Haddock 2004) , as well as Leptomedusae ( Harbison et al. 1977). Gasca and Browne (2017) provide additional records of associations with the salps Metcalfina hexagona ( Quoy & Gaimard, 1824) and Salpa maxima ( Forsskål, 1775) , the siphonophore Rosacea cymbiformis (Della Chiaje, 1830) and the ctenophore Cestum veneris Lesueur, 1813 . It has also been observed with the heteropod, Pterotrachea sp. ( Harbison et al. 1977), P. hippocampus Philippi, 1836 ( Gasca & Haddock 2004) and P. coronata Forsskål, 1775 ( Gasca & Browne 2017).
Distribution. Relatively common and widely distributed in the tropical and temperate regions of all the world’s oceans, including the Mediterranean Sea. In the Atlantic it ranges from about 60°N to 32°S. In the Indian Ocean it is common in the tropics and ranges from the Red Sea to south of Australia, to about 55°S ( Vinogradov 1962). In the Pacific it occurs generally below 40°N to south of New Zealand, to about 58°S ( Spence Bate 1862). Most records are from surface waters to 300– 400 m.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Brachyscelus crusculum Spence Bate, 1861
| Zeidler, Wolfgang 2021 |
Brachyscelus stebbingi
| Boone, L. 1935: 230 |
Brachyscelus mediterranea
| Senna, A. 1903: 5 |
Brachyscelus mediterraneus
| Chevreux, E. 1900: 154 |
Brachyscelus acuticaudatus Stebbing, 1888: 1555–1556
| Grice, G. D. & Hart, A. D. 1962: 300 |
| Stebbing, T. R. R. 1888: 1556 |
Brachyscelus crusculum
| Gasca, R. & Browne, W. E. 2017: 3 |
| Souza, C. S. & Conceicao, L. R. & Mafalda JR, P. O. 2016: 344 |
| Zeidler, W. 2016: 47 |
| Valencia, B. & Lavaniegos, B. & Giraldo, A. & Rodriguez-Rubio, E. 2013: 51 |
| Gasca, R. & Morales-Ramirez, A. 2012: 229 |
| Bucklin, A. & Ortman, B. D. & Jennings, R. M. & Nigro, L. M. & Sweetman, C. J. & Coply, N. J. & Sutton, T. & Wiebe, P. H. 2010: 2239 |
| Mori, M. & Suzuki, Y. & Yamaki, A. & Lindsay, D. J. 2010: 10 |
| Costa, F. & Krapp, T. & Ruffo, S. 2009: 103 |
| Gasca, R. 2009: 89 |
| Gasca, R. 2009: 66 |
| Gasca, R. & Manzanilla, H. & Suarez-Morales, E. 2009: 1497 |
| LeCroy, S. E. & Gasca, R. & Winfield, I. & Ortiz, M. & Escobar-Briones, E. 2009: 969 |
| Zeidler, W. & De Broyer, C. 2009: 63 |
| Gasca, R. & Franco-Gordo, C. 2008: 569 |
| Browne, W. E. & Haddock, S. H. D. & Martindale, M. Q. 2007: 819 |
| Gasca, R. 2007: 118 |
| Potier, M. & Marsac, F. & Cherel, Y. & Lucas, V. & Sabatie, R. & Maury, O. & Menard, F. 2007: 65 |
| Brusca, R. C. & Hendrickx, M. E. 2005: 151 |
| Gasca, R. & Haddock, S. H. D. 2004: 532 |
| Vinogradov, G. M. & Hernandez, F. & Tejera, E. & Leon, M. E. 2004: 15 |
| Gasca, R. 2003: 308 |
| Gasca, R. 2003: 118 |
| Gasca, R. & Shih, C-T. 2003: 95 |
| Escobar-Briones, E. & Winfield, I. & Ortiz, M. & Gasca, R. & Suarez, E. 2002: 367 |
| Lima, M. C. G. & Valentin, J. L. 2001: 473 |
| Lowry, J. K. 2000: 325 |
| Barkhatov, V. A. & Vinogradov, M. E. 1999: 808 |
| Vinogradov, G. M. 1999: 1145 |
| Montu, M. 1998: 599 |
| Zeidler, W. 1998: 96 |
| Lin, J. & Chen, M. & Chen, R. 1996: 231 |
| Lin, J. & Chen, M. & Chen, R. 1995: 118 |
| Shih, C. - T. & Chen, Q. - C. 1995: 178 |
| Tarling, G. A. & Ward, P. & Sheader, M. & Williams, J. A. & Symon, C. 1995: 34 |
| Lin, J. & Chen, R. 1994: 115 |
| Montu, M. 1994: 132 |
| De Broyer, C. & Jazdzewski, K. 1993: 118 |
| Vinogradov, G. M. 1993: 45 |
| Zeidler, W. 1992: 115 |
| Vinogradov, G. M. 1991: 261 |
| Zeidler, W. 1991: 133 |
| Young, J. W. 1989: 717 |
| Lin, J. & Chen, R. 1988: 325 |
| Young, J. W. & Anderson, D. T. 1987: 717 |
| Nagata, K. 1986: 274 |
| Brusca, G. J. 1981: 31 |
| Laval, P. 1980: 20 |
| Zeidler, W. 1978: 28 |
| Harbison, G. R. & Biggs, D. C. & Madin, L. P. 1977: 472 |
| Harbison, G. R. & Biggs, D. C. & Madin, L. P. 1977: 457 |
| Shulenberger, E. 1977: 379 |
| Tranter, H. A. 1977: 649 |
| Thurston, M. H. 1976: 388 |
| Brusca, G. J. 1973: 9 |
| Yoo, K. I. 1971: 43 |
| Dick, R. I. 1970: 68 |
| Hure, J. & Scotto di Carlo, B. & Basile, A. 1969: 603 |
| Hurley, D. E. 1969: 33 |
| Pillai, N. K. 1966: 225 |
| Sheard, K. 1965: 244 |
| Siegfried, W. R. 1963: 6 |
| Kane, J. E. 1962: 310 |
| Vinogradov, M. E. 1962: 23 |
| Evans, F. 1961: 200 |
| Hurley, D. E. 1960: 282 |
| Irie, H. 1957: 10 |
| Irie, H. 1957: 355 |
| Bulycheva, A. I. 1955: 1048 |
| Hurley, D. E. 1955: 181 |
| Reid, D. M. 1955: 25 |
| Irie, H. 1948: 36 |
| Shoemaker, C. R. 1948: 13 |
| Shoemaker, C. R. 1945: 242 |
| Dakin, W. J. & Colefax, A. N. 1940: 123 |
| Pirlot, J. M. 1939: 46 |
| Boone, L. 1935: 222 |
| Chevreux, E. 1935: 196 |
| Barnard, K. H. 1932: 292 |
| Barnard, K. H. 1930: 432 |
| Pirlot, J. M. 1930: 25 |
| Pirlot, J. M. 1929: 139 |
| Schellenberg, A. 1927: 649 |
| Spandl, H. 1927: 210 |
| Chevreux, E. & Fage, L. 1925: 427 |
| Stephensen, K. 1925: 172 |
| Spandl, H. 1924: 32 |
| Stewart, D. A. 1913: 262 |
| Sexton, E. W. 1911: 223 |
| Walker, A. O. 1909: 54 |
| Holmes, S. J. 1908: 490 |
| Tattersall, W. M. 1906: 26 |
| Chevreux, E. 1900: 153 |
| Norman, A. M. 1900: 134 |
| Chevreux, E. 1893: 70 |
| Stebbing, T. R. R. 1888: 1544 |
| Bovallius, C. 1887: 31 |
| Claus, C. 1876: 519 |
| Claus, C. 1873: 468 |
| Spence Bate, C. 1862: 333 |
| Spence Bate, C. 1861: 10 |