
Cephalozygoptera Archibald, Cannings & Erickson
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4934.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:79895443-4597-42A5-AF8A-023EACB20E10 |
|
DOI |
https://doi.org/10.5281/zenodo.4672713 |
|
persistent identifier |
https://treatment.plazi.org/id/03B487C2-0008-FF88-FF5B-FA67FD3D1194 |
|
treatment provided by |
Plazi |
|
scientific name |
Cephalozygoptera Archibald, Cannings & Erickson |
| status |
subord. nov. |
Cephalozygoptera Archibald, Cannings & Erickson , new suborder
To expand the Zygoptera concept to include the head and eye morphologies of the Dysagrionidae and Sieblosiidae (see below) would result in the loss of these few primary, unambiguous, and easily observable diagnostic character states defining the suborder. We, therefore, group these families and possibly the Whetwhetaksidae , whose wings indicate a close relationship with them, as the new suborder Cephalozygoptera . If a future fossil (presumably in amber, as this is highly unlikely to be seen in any compression fossil) shows that the ligula differs from that of Zygoptera , this may be considered a synapomorphy of the Cephalozygoptera ; if it is consistent with the Zygoptera , it would be a synapomorphy of Zygoptera + Cephalozygoptera .
Diagnosis. Cephalozygoptera are odonates most like Zygoptera by a combination of: fore- and hind wings petiolate, with similar shape and venation including quadrangles that are closed and not crossed, CuA simple, and a well-developed nodus; an oblique thorax; gracile body; male clasping anal appendages comprised of a pair of dorsal cerci (without paddle-shaped extensions) and a pair of well-developed ventral paraprocts; and ocelli arranged in an equilateral triangle; but may be easily distinguished from them by any of:
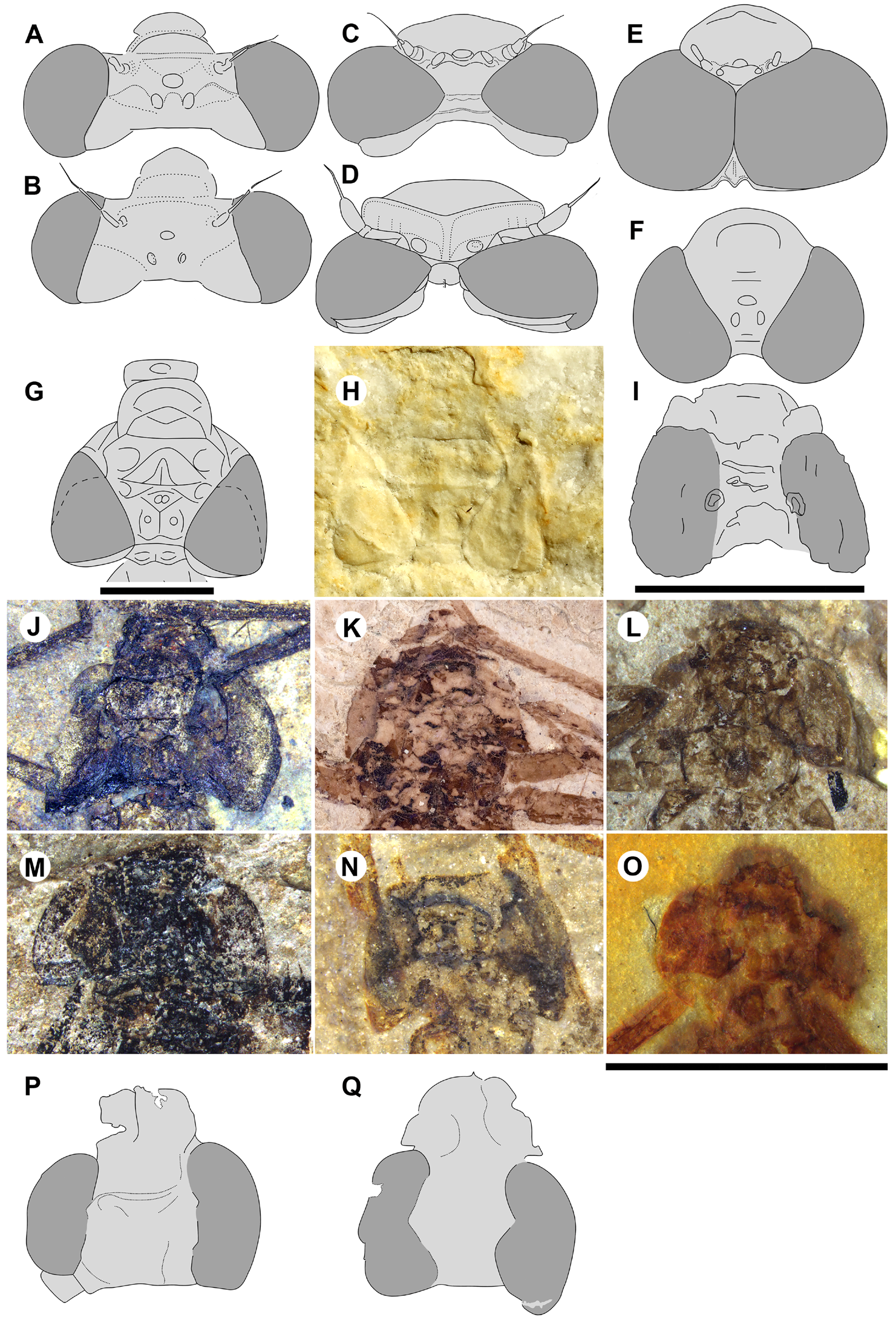
1- head width across eyes about twice the length from anterior margin of antefrons to posterior of occiput; [ Zygoptera : usually about three to five times as wide]; and compound eyes that are:
2- more or less adpressed to head, posterolateral corners protruding posteriorly to varying degrees, more or less rounded, sometimes acutely [ Zygoptera : distinctly bulging laterally from head capsule];
3- separated by about one eye’s width dorsally (ratio of width between eyes/width eye has a range of 0.8–1.9 and a mean 1.0), n=11 ( Table 1 View TABLE 1 ) [ Zygoptera : usually more than twice an eye’s width].
Included taxa. The families Dysagrionidae and Sieblosiidae , and possibly the Whetwhetaksidae , new family.
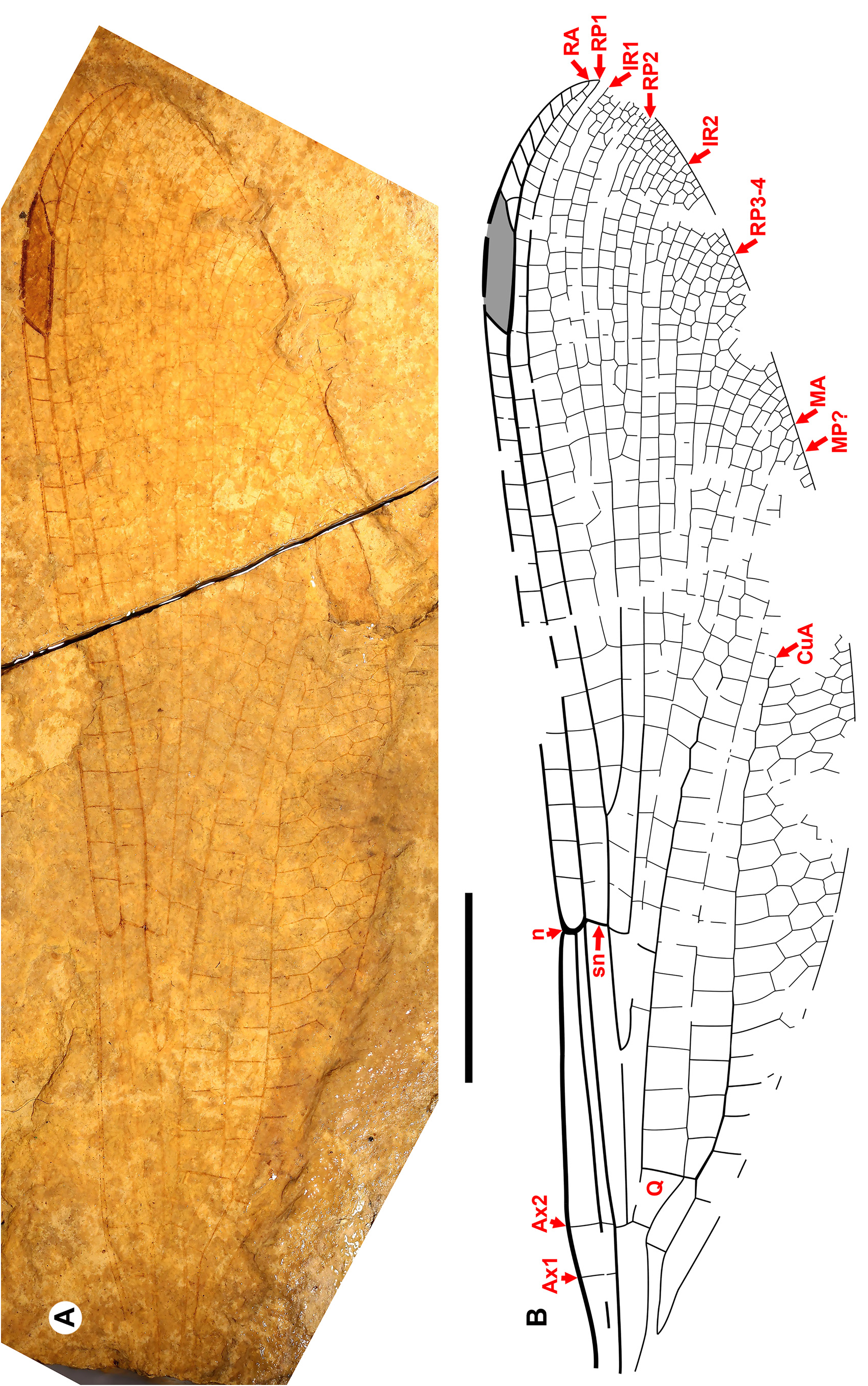
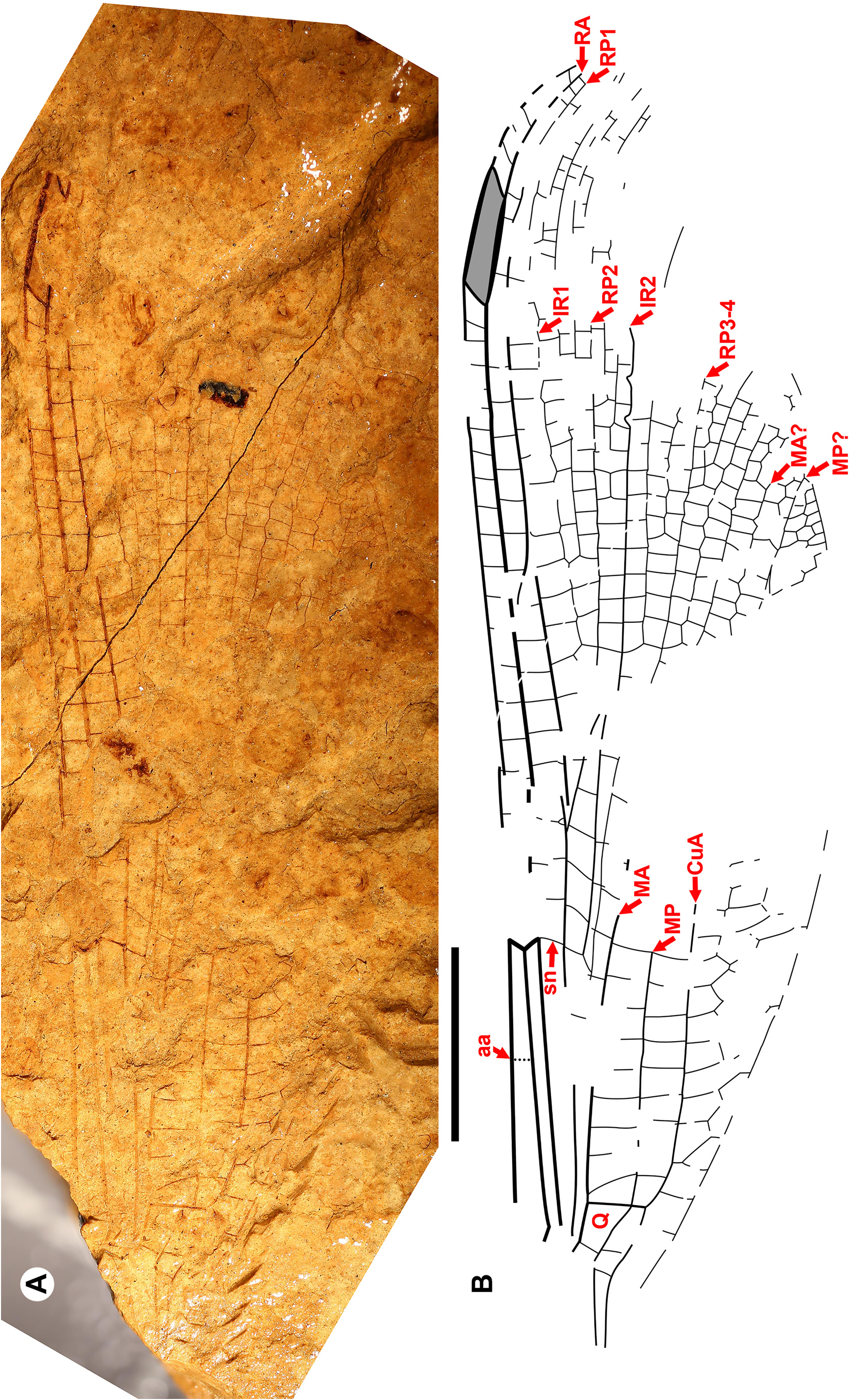
Description. With character states of the diagnosis and further of the wings: Ax0 present; nodus at about a quarter wing length or more; anterior anal vein separates at a right angle from the posterior anal vein briefly before joining CuP (i. e., is briefly free distal to petiole); RP3-4 origin (midfork) in middle third between arculus and subnodus; antesubnodal space without crossveins; distinctive quadrangle (closed, distal side longer than proximal, posterior longer than anterior); long to very long pterostigmata (2.5 to about 10 times width if the Whetwhetaksidae is a member).
Etymology. The name Cephalozygoptera is derived from the Greek κεφάλή, kefáli, “head”, and Zygoptera (ζυγός, zugós, even + πτερόν, pterón, wing, referring to the similar-shaped fore- and hind wings), indicating that they are distinguished from damselflies by their heads.
Discussion. Sieblosiidae and Whetwhetaksidae . Handlirsch (1907) excluded Sieblosia jucunda from the Zygoptera , believing that it belongs to the Anisozygoptera without explanation. This view was later rejected by Fischer (1974) and Nel et al. (1993), who restored it to the Zygoptera . The type specimen (apparently lost: Fischer 1974) was rather complete, including the head, which Hagen noted is almost as wide as long. In his drawing ( Hagen 1858: his Plate 24, Fig. 1 View FIGURE 1 ), although lacking detail, the head closely matches those of the Dysagrionidae (reproduced here as our Fig. 10 I View FIGURE 10 ). He assumed, however, that it was compressed post mortem (page 121: “kopf platt gedrückt”). Fleck et al. (2004) recognised that the head of Sieblosiidae is not transversely elongate with the compound eyes placed far apart. They further found that their wings do not share any unambiguous apomorphy with the Zygoptera and that particularly with their nodal morphology and curved CuP they may not belong to the suborder. Following this, Bechly (2015) treated the family as “ Anisozygoptera ”.
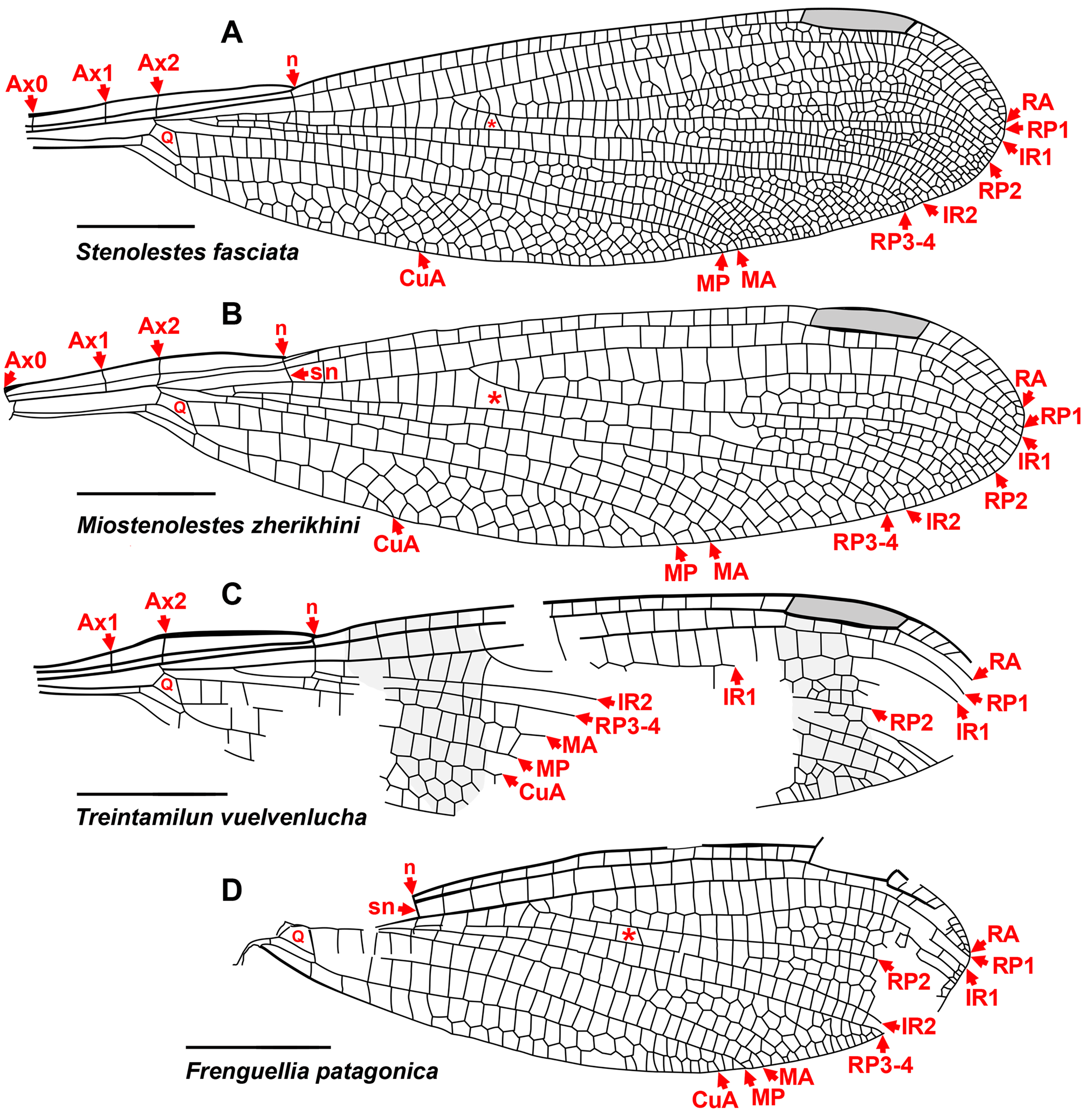
The wings of Sieblosiidae and Dysagrionidae have strong similarities ( Nel et al. 2005a) ( Fig. 12 View FIGURE 12 ). They share with Dysagrionidae character states 2 through 7 of its diagnosis, notably the distinctive quadrangle shape, character state 3. The wings of both are broad, with a short petiole and bear a long pterostigma. Crossvein Ax0 has been found in numerous species, see Germanostenolestes lutzi (2012, their Fig. 3 View FIGURE 3 ), Stenolestes fischeri Nel , holotype MNHN- LP-R.06677 ( Nel et al. 1997, their Fig. 6 View FIGURE 6 ), and in Stenolestes fasciata Nel et al. ( Nel et al. 2005a, their Fig. 9 View FIGURE 9 ), Stenolestes cerestensis Nel et al. ( Nel et al. 2005a, their Fig. 14 View FIGURE 14 ), and Miostenolestes zherikhini ( Nel et al. 2005a, their Fig. 5 View FIGURE 5 ). As in the Dysagrionidae , the wing base of most published Sieblosiidae fossils where Ax0 might be found is missing or damaged, and so this represents a minimum of specimens where this may be present; judging from published drawings and photographs, none may be excluded as possessing it. As in Petrolestinae but not the Dysagrioninae , the origin of IR2 is in the middle third between the arculus and nodus, close to that of RP3-4. Nel & Paicheler (1994) suggested that Sieblosiidae and Dysagrionidae might be an example of convergent evolution.
Sieblosiidae differ from the above diagnosis of the Dysagrionidae by character state 8, width of the CuA–A space, which is usually two or more cells wide as in Dysagrionidae , but may be one cell wide (e.g., see Fig. 12B View FIGURE 12 , Miostenolestes zherikhini ); by 9, CuA terminates on the margin proximal to the mid-wing; and 1, crossvein O is present. The nodus of Sieblosiidae differs as well by ScP passing through it and terminating on the anterior margin at an oblique angle. Although the pterostigmata of Dysagrionidae are long, those of the Sieblosiidae are longer.
The new family Whetwhetaksidae is only known by wings, which share character states 1 and 3 through 9 of the Dysagrionidae diagnosis and their general shape is like that of the Dysagrionidae and Sieblosiidae : broad, with a short petiole. The origin of IR2 is in the middle third between the arculus and nodus as in Sieblosiidae and the Petrolestinae, but not the Dysagrioninae .
These wings are notably distinct, however, as the arculus is in a proximal position near Ax1 as is found in some odonates outside of the Zygoptera . This feature is unknown in extant Zygoptera except in the Amazonian Heliocharis amazona Selys (Calopterygoidea, Dicteriadidae ), and is present in two Paleogene Euphaeidae (the Priabonian Litheuphaea ludwigi Bechly and the Chattian Parazacallites aquisextanea Nel : Bechly 1998; Nel 1988). Whetwhetaksidae possess an extremely long pterostigma, unknown in Zygoptera , but like those found in the anisopteran Petaluridae . It is up to about ten times longer than wide, and is about a quarter to almost a third the length of the nodus to the wing apex. Further, the nodus is distant from the wing base, at about 38–40% wing length.
Groups with similar wings that are excluded. The Frenguelliidae are odonates from the Ypresian and Lutetian of Patagonia, Argentina, known only from wings ( Fig. 12C, D View FIGURE 12 ). They are considered by some authors not to belong to the Zygoptera by their curved CuP ( Petrulevičius & Nel 2003, 2007, 2013; Petrulevičius 2017, 2019). The wing of Treintamilun vuelvenlucha Petrulevičius resembles those of the Dysagrionidae in many aspects, but differs by its vertical subnodus and—importantly—its curved CuP. Those of Nelala chori Petrulevičius lack the characteristic Cephalozygoptera quadrangle shape. The suborder affinity of Frenguelliidae is unclear, and we follow Petrulevičius (2017, 2019) and Petrulevičius & Nel (2003, 2007, 2013) in treating it as Odonata suborder indet., pending more complete specimens.
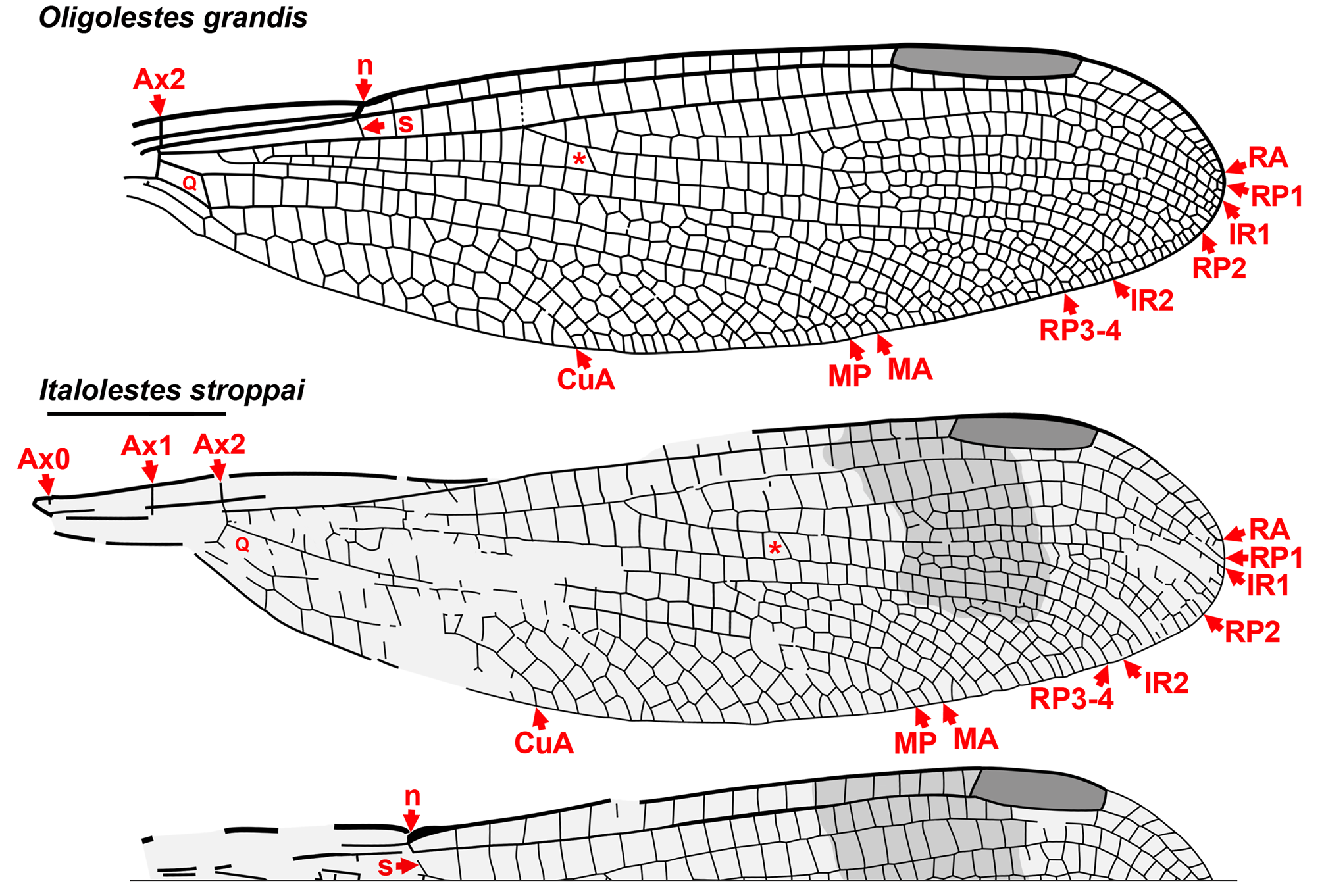
The wings of Oligolestes grandis (Statz) from the Chattian of Rott, Germany and Italolestes stroppai Nel et al. from the Messinian (late Miocene) of Italy ( Gentilini & Bagli 2004; Nel et al. 2005 a, 2005c) share many traits with the Sieblosiidae ( Fig. 13 View FIGURE 13 ) but differ in important ways, e.g., the quadrangle of Oligolestes Schmidt is somewhat narrower and the bases of RP3-4 and IR2 are in a distinctly different position. Both genera differ significantly from the Sieblosiidae in possessing a subnodus of normal obliquity and ScP not passing through the nodus (see Nel et al. 2005a). We agree with Nel et al. (2005a) that they might be best treated as of uncertain position pending the discovery of more complete fossils.
Cephalozygoptera terminal appendages. Only the male terminal appendages in Cephalozygoptera are known, unless those of the female are present on some Sieblosiidae fossils of which we are not aware. The well-developed cerci and paraprocts at the apex of the abdomen are used in extant Zygoptera to grasp the female during mating. As the terminal appendages of male Cephalozygoptera are so like those of Zygoptera , we predict that their female genitalia will be found to be correspondingly similar. We are not aware of any Cephalozygoptera fossil that preserves secondary male genitalia.
Cladistic analyses. Searches performed using morphological data (Appendix C) recovered a single most-parsimonious tree (MPT). Analysis 1, of the relationships of the suborders of Odonata, Burmese amber genera not assigned to suborder, and Tarsophlebiidae , produced one MPT ( Fig. 14 View FIGURE 14 ). Constraining Tarsophlebiidae as the outgroup recovers Anisoptera and Anisozygoptera as sister taxa with high support, consistent with other recent analyses ( Westfall et al. 2014 and references therein). Cephalozygoptera and Zygoptera are recovered as sister taxa with high support, while Burmadysagrion , Electrodysagrion , and Paleodysagrion, formerly considered to belong to the Dysagrionidae , are recovered as members of Zygoptera with moderate to high support. Zygoptera , therefore, was used as an outgroup in Analyses 2 and 3.
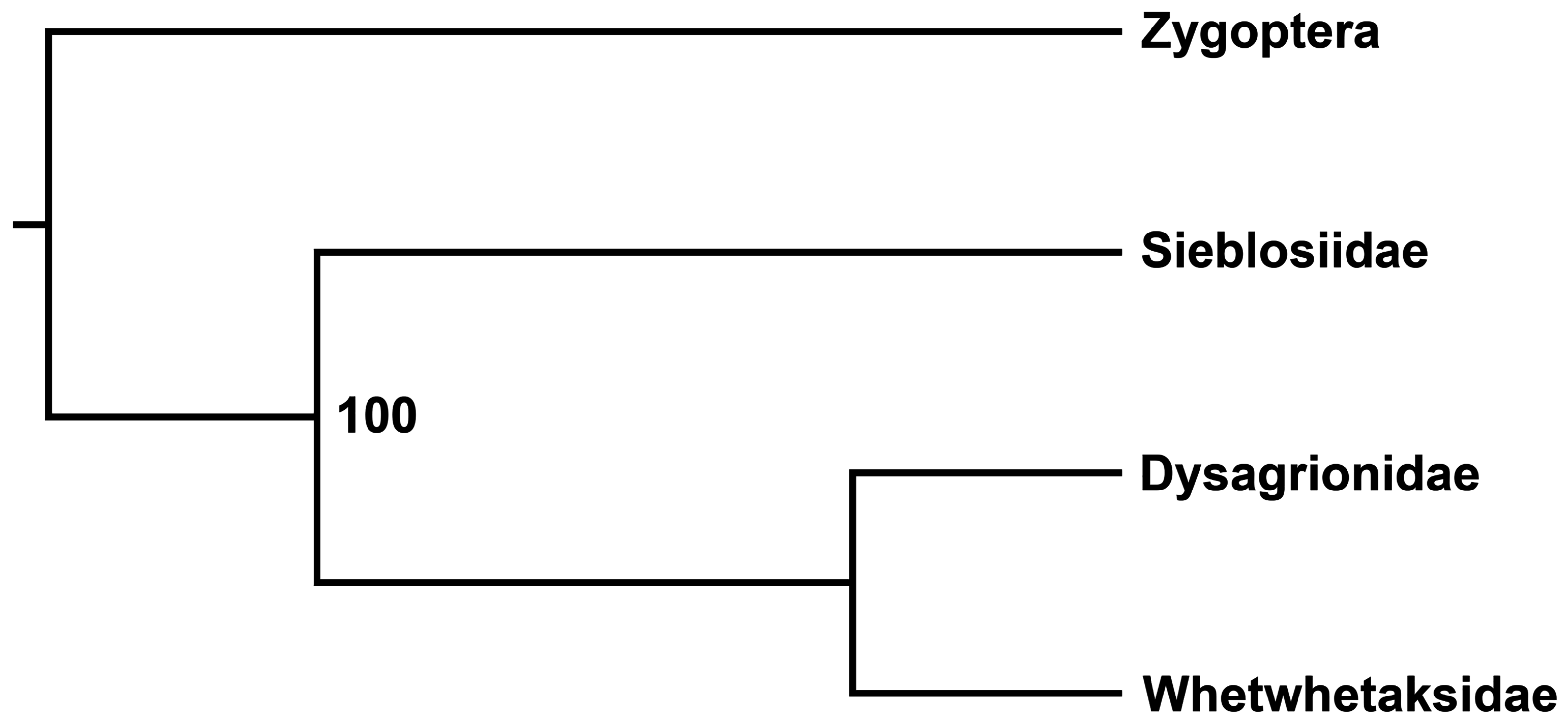
In Analysis 2, the relationship of Whetwhetaksidae to Cephalozygoptera and its families, produced a single MPT ( Fig. 15 View FIGURE 15 ) that provides weak support for Dysagrionidae and Whetwhetaksidae as sister taxa.
Analysis 3, the internal phylogeny of Dysagrionidae , is discussed below.
History and biogeography of the Cephalozygoptera . The Cephalozygoptera has been found in localities across the Northern Hemisphere ( Table 3 View TABLE 3 ) from the Aptian of China; possibly the Thanetian of France ( Valerea , Thanetophilosina ) and the Thanetian or Ypresian of Alaska, United States of America ( Garrouste & Nel 2019: unnamed); the Ypresian of Denmark, Canada, and the United States of America; the Lutetian of Germany; the Priabonian of Colorado, United States of America, the United Kingdom, and Russia (European and Pacific); the Rupelian (early Oligocene) of Russia (Caucasus) and Germany; Chattian (late Oligocene) of France; Langhian/Serravallian (middle Miocene) of Russia (Caucasus) and Bulgaria; Serravallian (middle Miocene) of Croatia and Germany; Tortonian (late Miocene) of Spain and France.
Bechly (pers. comm.) reports a specimen from the Ypresian Green River Formation that bears the distinctive Cephalozygoptera head morphology, but is in ways quite different from the families that we include, suggesting that the suborder may have been larger and more diverse than we understand it here.
The Dysagrionidae dominanted the Eocene Cephalozygoptera across the Holarctic (see Table 3 View TABLE 3 and references therein). They were by far the dominant Odonata in more temperate far-western montane North America, the Okanagan Highlands (in upper microthermal mean annual temperatures, i.e. 13°C and below: Wolfe & Wehr 1987; Archibald & Farrell 2003; Greenwood et al. 2005; Archibald et al. 2014), with 70 fossils in seventeen species assigned to four genera and one genus-level parataxon genus, contrasted with seven specimens of the single species of Whetwhetaksidae ; nine fossil dragonflies there in two families, seven genera and eight species; and one undescribed fossil damselfly wing. It is unclear if the larger wings of dragonflies depress their fossil abundance relative to their community abundance by taphonomic bias through increased floating time before deposition on the substrate ( Martínez-Delclòs & Martinell 1993, Wagner et al. 1996).
They were less common, but still well-represented in the hot lowlands of mid-continental Ypresian Colorado and Wyoming (mean annual temperatures upper mesothermal to megathermal, megathermal = 20°C and over: Archibald et al. 2011b), and are fewer yet in the upland Priabonian of Colorado (likely similar mean annual temperature as the Okanagan Highlands: Allena et al. 2020), where they were mixed with a variety of dragonflies and damselflies, and were present to lesser degrees in the Eocene of Europe and Asia.
After the Eocene, the Dysagrionidae become rare, with one ( Primorilestes magnificus ), perhaps two ( Phenacolestes ? coloratus) occurrences in the Miocene. Sieblosiidae dominate the Cephalozygoptera in the Oligocene and Miocene, the globally cooler post-Eocene icehouse world climatic regime ( Zachos et al. 2001), restricted to the Palearctic Realm. Their last record, and so that of the suborder, is in the late Miocene.
The cool, higher elevations of the Okanagan Highlands is where we see many of the earliest occurrences of the floral taxa and community associations of modern temperate Northern Hemisphere forests that later descended to lower elevations and spread throughout and predominate in much of the Holarctic as temperature seasonality increased outside of the tropics bringing colder winters, the latitudinal gradient of mean annual temperature steepened, and cooler global temperatures were established following the close of the Eocene (e.g., Graham 1999; Zachos et al. 2001, 2008). As the Cephalozygoptera genera Okanagrion and Okanopteryx were the dominant Odonata of the montane Okanagan Highlands forests, why did they not spread with them and flourish across the Holarctic to the present? There is no obvious explanation.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.