
Okanagrion Archibald & Cannings, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4934.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:79895443-4597-42A5-AF8A-023EACB20E10 |
|
DOI |
https://doi.org/10.5281/zenodo.4672736 |
|
persistent identifier |
https://treatment.plazi.org/id/03B487C2-003A-FFBD-FF5B-FD33FDE014A8 |
|
treatment provided by |
Plazi |
|
scientific name |
Okanagrion Archibald & Cannings |
| status |
gen. nov. |
Okanagrion Archibald & Cannings , new genus
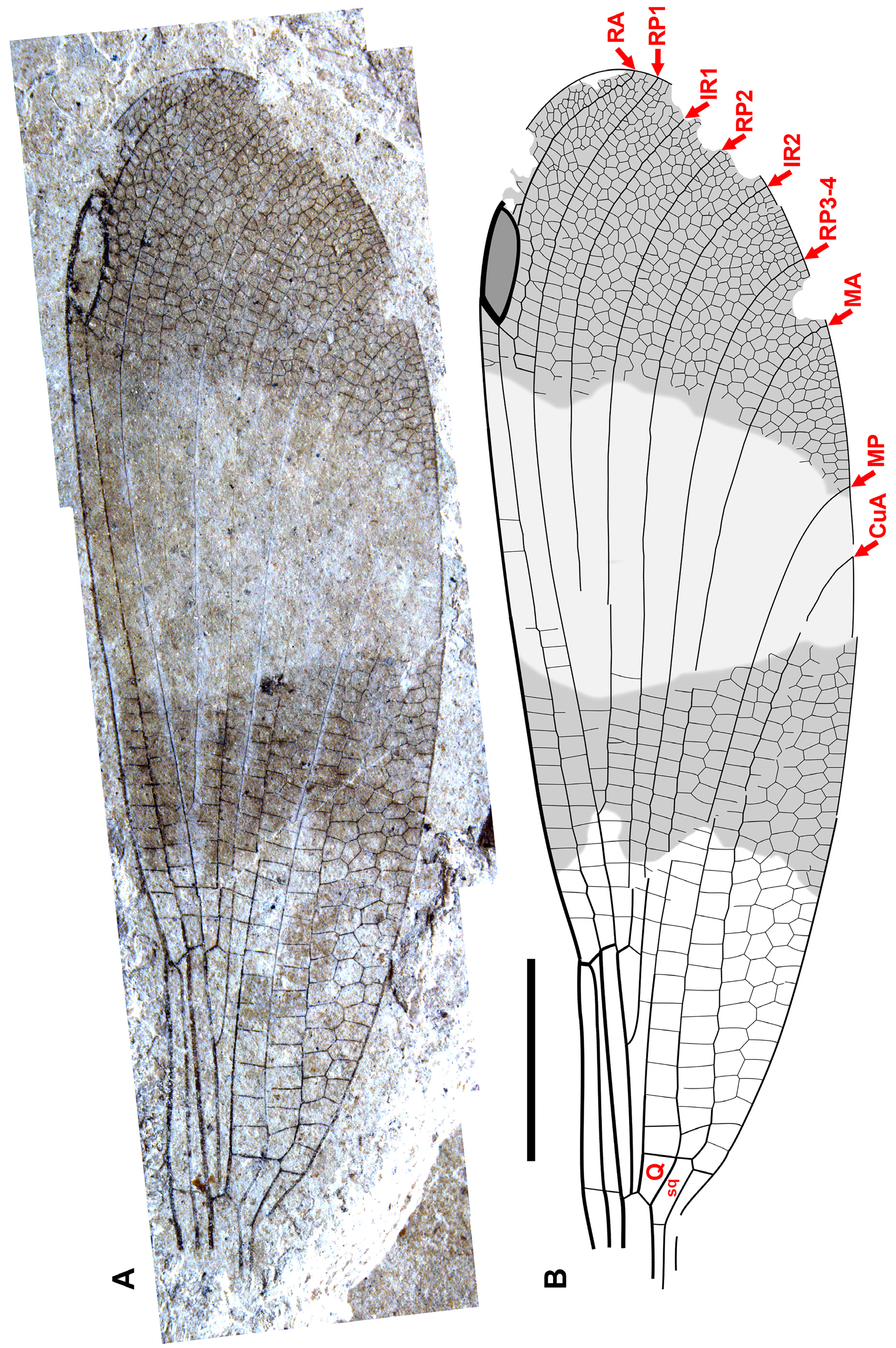
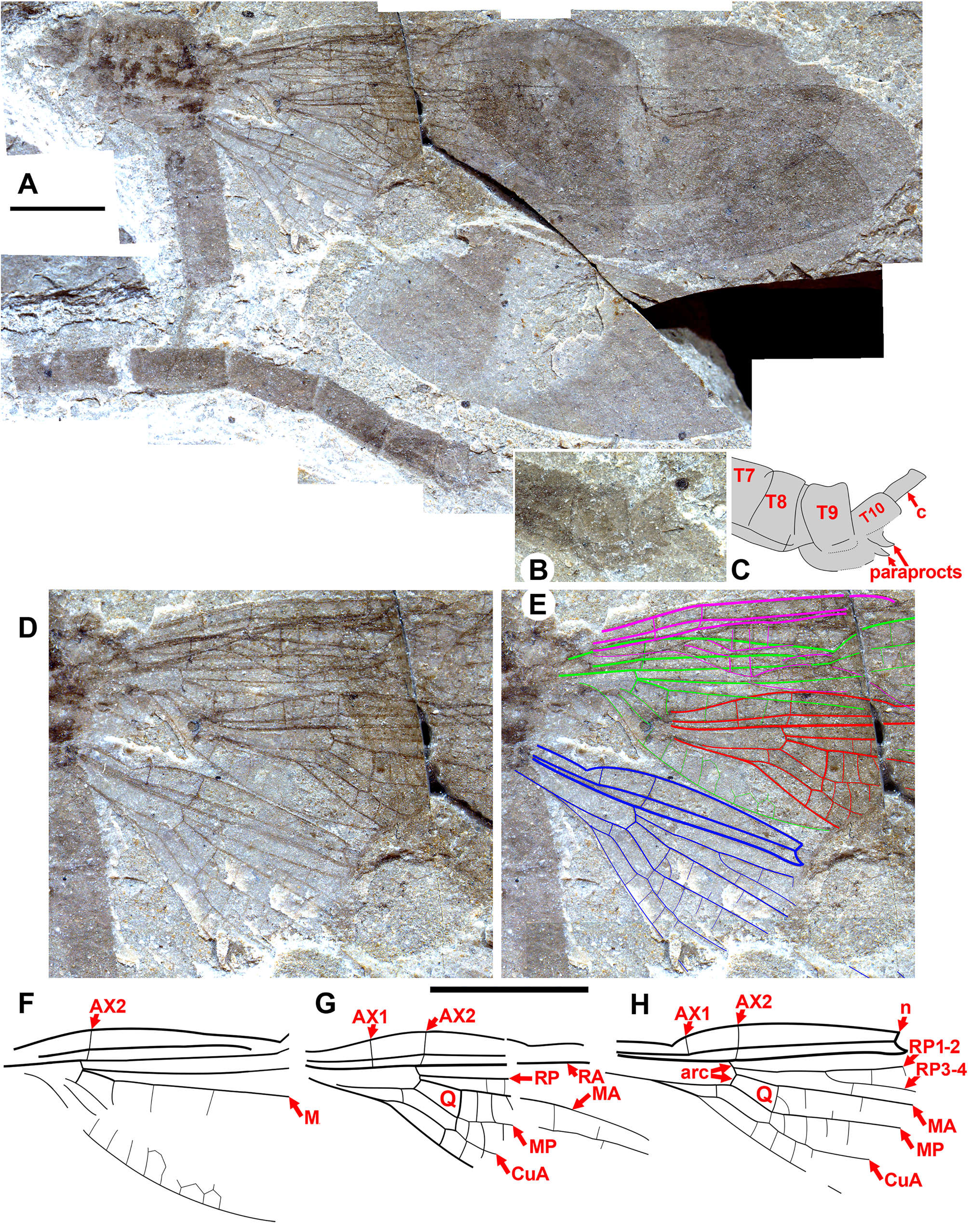
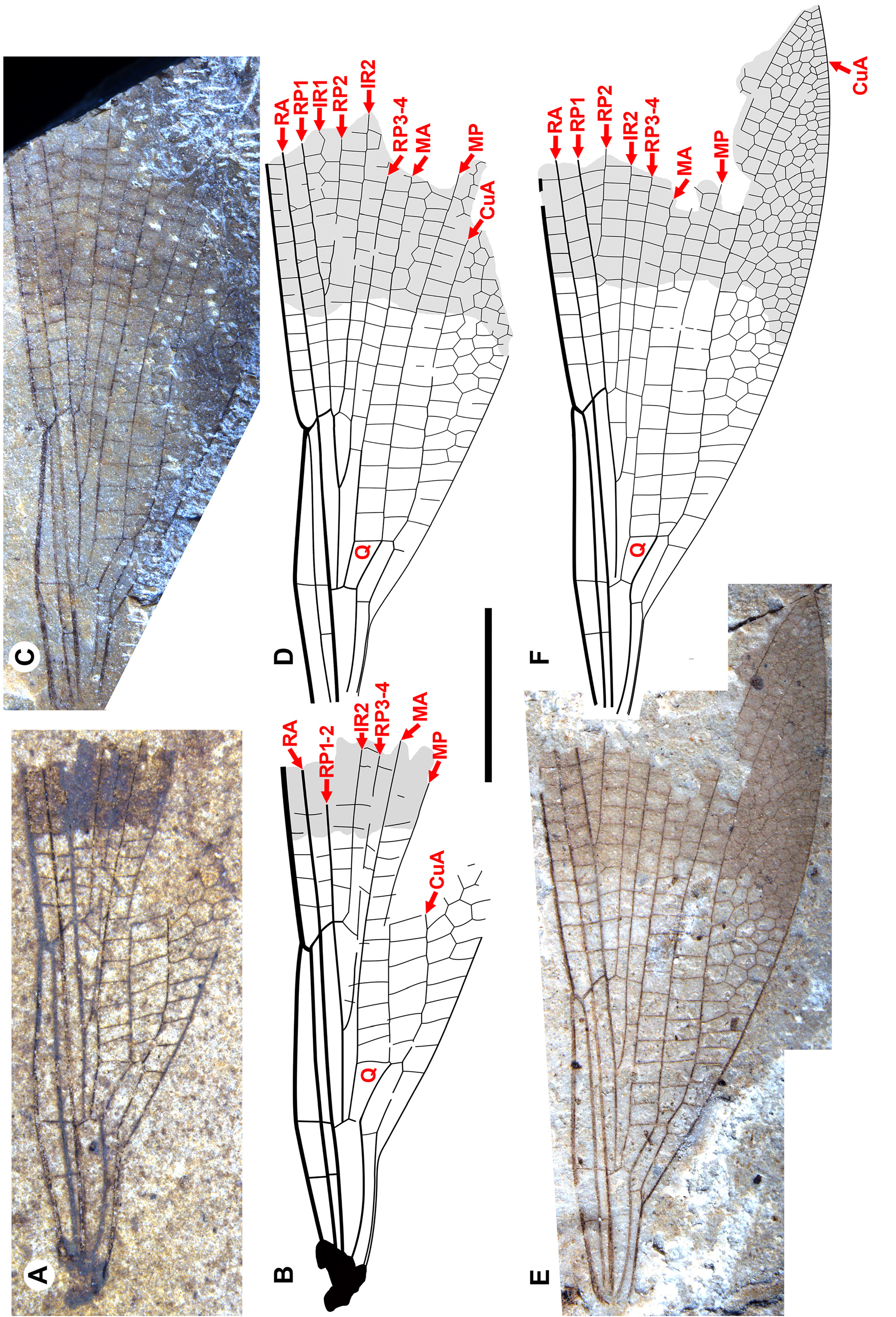
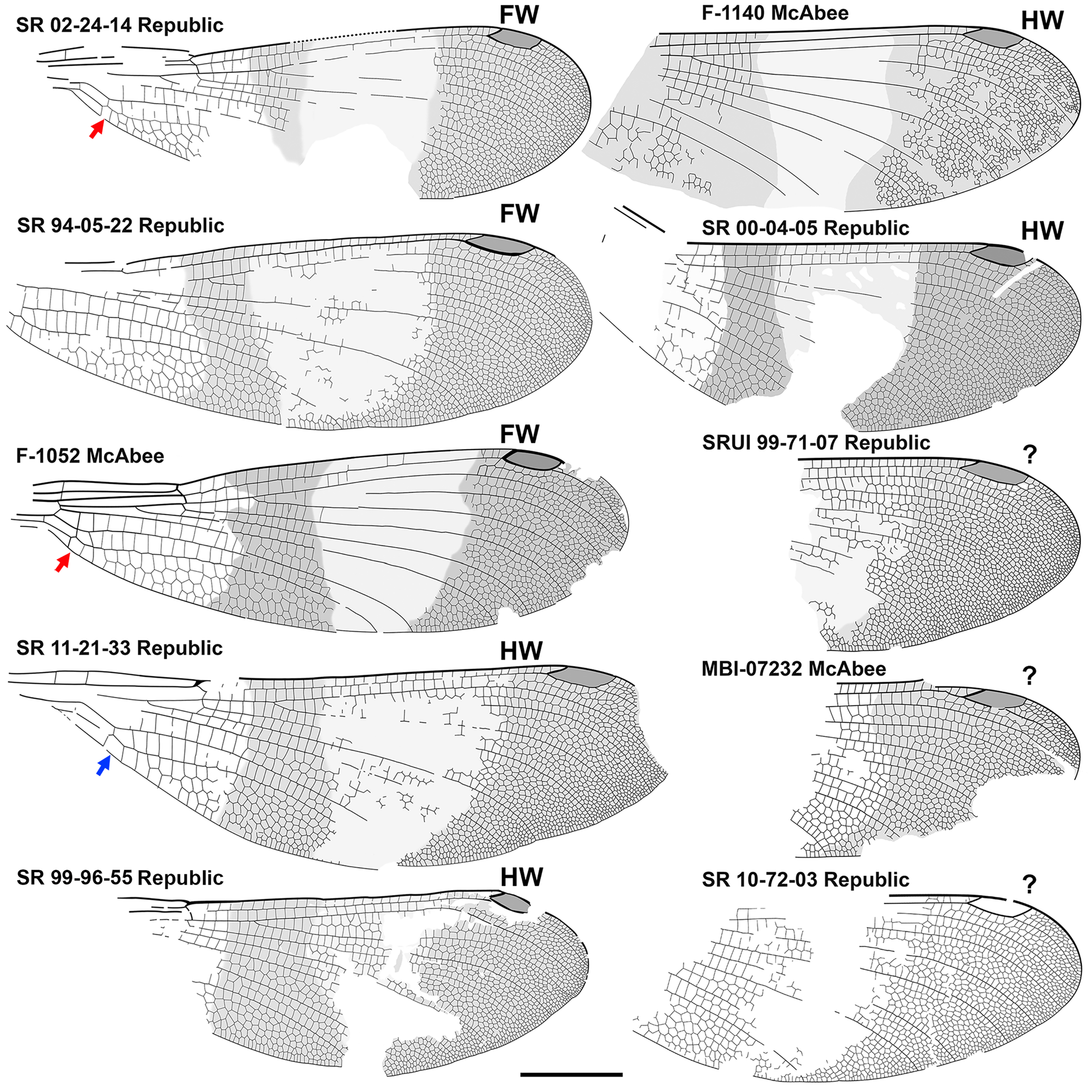
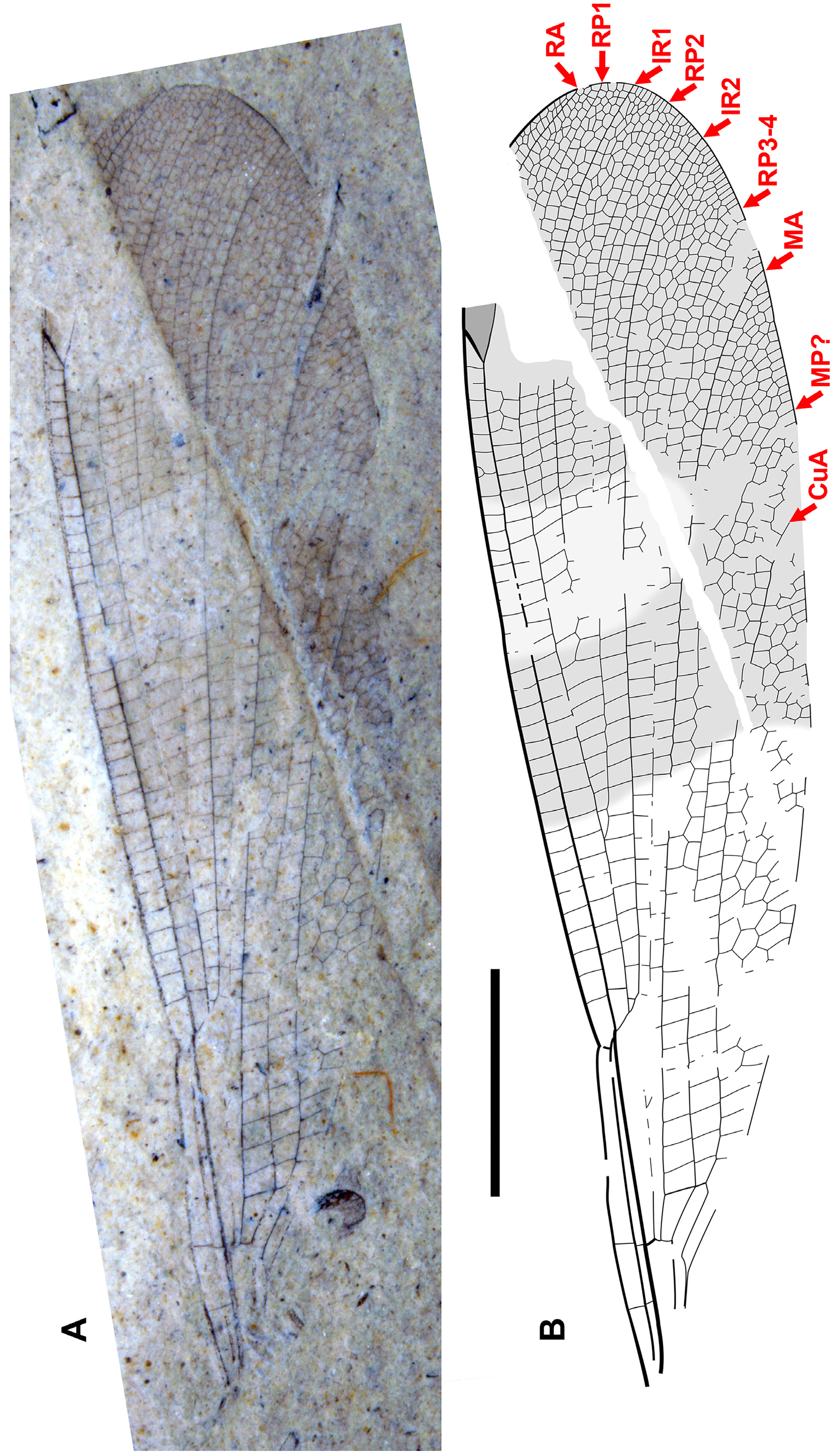
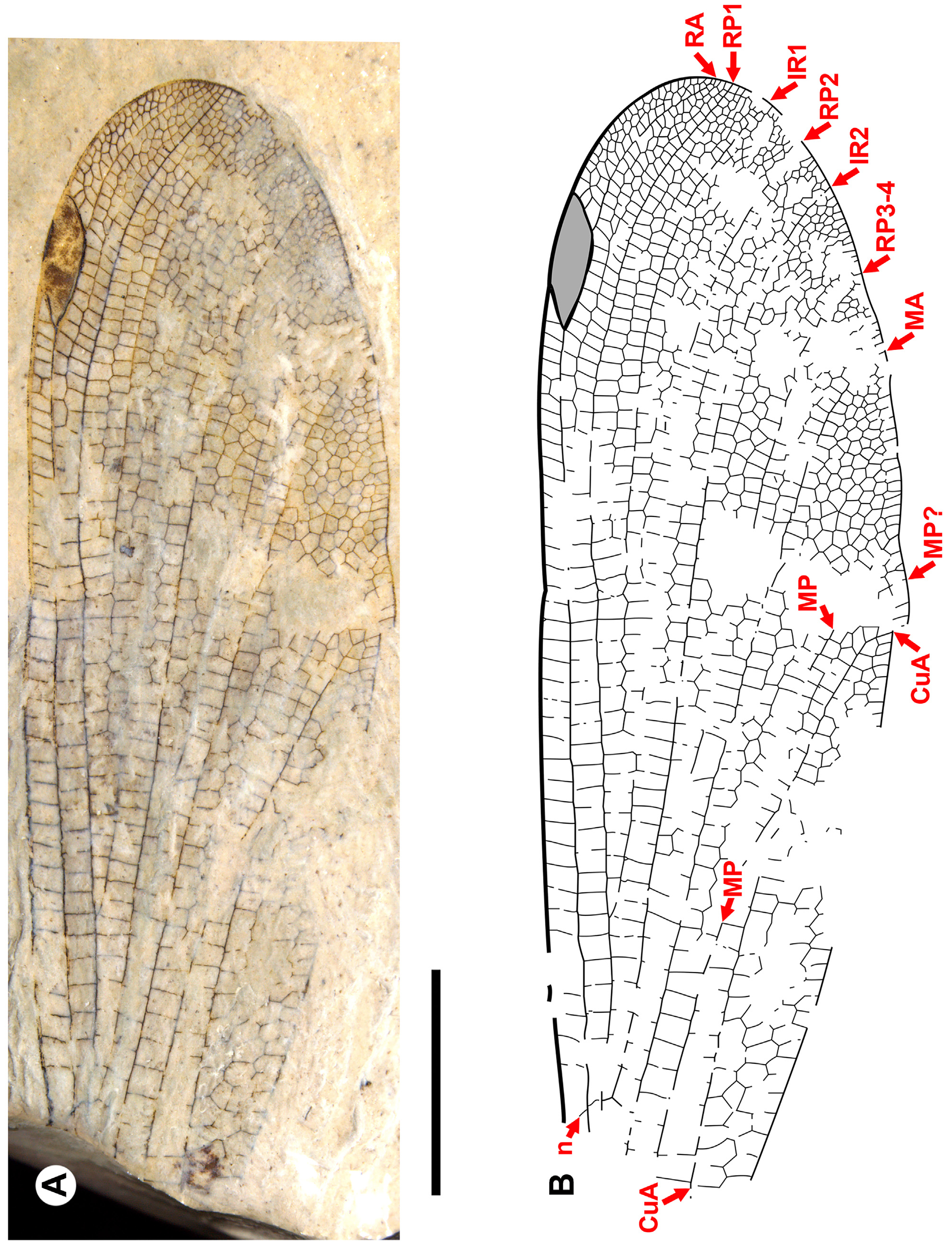
Figs. 19–43 View FIGURE 19 View FIGURE 20 View FIGURE 21 View FIGURE 22 View FIGURE 23 View FIGURE 24 View FIGURE 25 View FIGURE 26 View FIGURE 27 View FIGURE 28 View FIGURE 29 View FIGURE 30 View FIGURE 31 View FIGURE 32 View FIGURE 33 View FIGURE 34 View FIGURE 35 View FIGURE 36 View FIGURE 37 View FIGURE 38 View FIGURE 39 View FIGURE 40 View FIGURE 41 View FIGURE 42 View FIGURE 43 .
Diagnosis. Wings distinct from all other genera of Dysagrioninae by: pterostigma 3–3.5, rarely 4 times longer than wide [separated from Okanopteryx : 2.5 times; Stenodiafanus : 4; Phenacolestes : 5; not separated from Furagrion : 3; Primorilestes : 3.5–4; Dysagrion : 4–7.5; Dysagrionites species: unknown]; no oblique brace vein [separated from Dysagrion , Furagrion, Okanoptyeryx , Stenodiafanus ; wing region not preserved in Electrophenacolestes , Dysagrionites species; not separated from Phenacolestes , Primorilestes ]; costal space 3–5 cells wide distal to pterostigma [distinct from Primorilestes , Okanopteryx , Stenodiafanus : all 2; Dysagrion , Furagrion , Phenacolestes : all 1; Electrophenacolestes , Dysagrionites species: not known]; subnodus with normal obliquity (reverse in one specimen of Okanagrion hobani , paratype 1, which we consider adventitious) [separated from Dysagrion , reversed obliquity; all others: normal obliquity, slight to notable]; no accessory antenodal crossveins [distinguished from Phenacolestes , Electrophenacolestes , some Dysagrion species: present; not separated from all others: also none]; IR2 origin rather evenly between RP1-2, RP3-4, connected to RP1-2, RP3-4 at similar level (some species IR2 origin close to or on RP1-2) [ Dysagrion rather evenly between RP1-2, RP3-4, but connected with RP1-2 distinctly more distally than to RP3-4; other genera IR2 origin rather evenly between RP1-2, RP3-4, connected to RP1-2, RP3-4 at similar level]; MA linear from quadrangle to margin [separated from Dysagrion: MA linear from quadrangle to zigzagged roughly mid-wing or further, linear again near margin; Furagrion : similar to Dysagrion but deeply curved in distal portion; Phenacolestes : very slightly zigzagged or not in mid-portion; Okanopteryx , Stenodiafanus , Dysagrionites species (known portions, none known to margin): linear to mid-wing, somewhat to strongly zigzagged to margin; not separated from Primorilestes : linear]; MA, MP far apart at margin [distinct from Dysagrion: MA, MP close, 1–2 cells apart at margin; not clearly distinct from Okanopteryx , Stenodiafanus as difficult to be sure of identity of MA near margin by strong zigzag, but close, probably separated by two cells; Dysagrionites species also difficult, perhaps one cell; not separated from Furagrion , Primorilestes , Electrophenacolestes: MA, MP widely separated]; CuA not zigzagged (or only very slightly so), linear to margin [distinct from Dysagrion : CuA increasingly zigzagged distal to level of nodus; Furagrion : slightly zigzagged distal to level of origin of RP2; Phenacolestes : may be very slightly zigzagged in parts; Okanoptyeryx, Stenodiafanus : zigzagged to various degrees distal to about mid-wing; not clearly distinct from Electrophenacolestes , Primorilestes : linear or very slightly zigzagged]; CuA–A space 4–7 cells wide [separated from Furagrion : 2; Stenodiafanus : 2 (most likely, wing incomplete); Dysagrionites delinei : 3; Dysagrionites sp. A: 2; Dysagrionites sp. B: probably 3 but unclear; not separated from Dysagrion : 4; Phenacolestes : 4; Primorilestes : 3–5; Electrophenacolestes : 4 (one column 5); Okanopteryx : 2–4]; postnodal, postsubnodal crossveins not aligned at least in proximal half [separated from Stenodiafanus , Okanopteryx , Primorilestes violetae ; Electrophenacolestes ; Furagrion : all aligned or somewhat so in proximal half; not distinct from Phenacolestes , Primorilestes madseni , Dysagrion : all not]; further from Dysagrion by nodus at 25–30% wing length [ Dysagrion : Ξ 35%]; further from Primorilestes by no intercalary vein in CuA–A space. Separated from Valerea (tentative Dysagrioninae ) by no linear supplementary sector between RP1, IR1 [ Valerea : with such sector]; origins of IR1, RP2 further from nodus; origin of IR2 between RP1-2, RP3-4 or near or on RP1-2 [IR2 origin not preserved, only preserved to level of nodus, where it is closer to RP3-4]. Most easily separated from genera regarded here as possible Dysagrionidae as follows: from Thanetophilosina by pterostigma 3–3.5, rarely 4 times longer than wide [about 6 times width], by no linear supplementary sectors between RP1, IR1, between IR1, RP2 [ Thanetophilosina : with such sectors]; from Chickaloon specimen by MA linear [beginning to zigzag in distal-most portion preserved]; from NHMUK I.9866/I.9718 by origin of RP3-4 [distinctly closer to subnodus]; from cf. Dysagrionidae genus A, species A (“ Megapodagrionidae ” genus A, species A of Petrulevičius et al. 2008) by width of CuA–A space [two cells wide, preserved portion, likely maximum], by crossvenation in apical third [not as dense, e.g., costal space apical to pterostigma two cells wide, RA–RP1, RP1–IR1 spaces one cell wide to apex]; no brace vein [present].
Type and included species. Type species: Okanagrion threadgillae new species; included: O. hobani , O. beardi , O. lochmum new species, O. angustum new species, O. dorrellae new species, O. liquetoalatum new species, and O. worleyae new species.
Description. As in diagnosis and the following. Colouration of wings varies: broad regions of dark or light fasciae or may be completely hyaline. Pterostigma 3–3.5 (rarely four) times longer than wide; four–seven cells subtend pterostigma (usually six); no accessory crossvein proximal to or distal to CuP; no well-defined supplementary sector between IR1, RP2; CuP proximal to arculus, between levels of Ax1, Ax2; petiole ends proximal to CuA, proximal to CuP; no oblique brace vein; postnodal, postsubnodal crossveins not aligned; CuA–A space maximum four–seven cells wide; origin of RP3-4 proximal to subnodus; no crossvein O; origin of IR2 at (or immediately proximal to) subnodus; IR1 zig-zagged in proximal half, CuA may be slightly zigzagged briefly about nodal level, apart from this all major veins linear (except MP zigzagged distally in O. dorrellae ), well defined; usually one–two hyperstigmal crossveins, less often none; two antenodal crossveins (Ax1, 2), rarely Ax0, no accessory antenodal crossveins; Ax2 at or very close to, immediately distal to arculus; CuA long, terminating on posterior margin beyond half length of arculus to apex.
Etymology. The genus name is a toponym derived from “Okanagan Highlands” and - agrion, the Greek ἄγρΙΟς, often used as a suffix in forming damselfly genus names. Gender neuter.
Range and age. McAbee, BC, Canada and the Klondike Mountain Formation at Republic, Washington, USA; second half of the Ypresian.
Discussion. Valerea and the Chickaloon specimen. Okanagrion wings share distinctive character states with that of the Thanetian Valerea multicellulata from Menat, France ( Garrouste et al. 2017). That insect has a notably large number of small cells in the costal and RA–RP1 spaces distal to the pterostigma, up to five cells wide, a condition that is only approached in the RA–RP1 space by some Mesozoic Aeschnidiidae , in which it is up to three cells wide ( Fleck & Nel 2003; Garrouste et al. 2017). In Okanagrion , there are usually three, and sometimes four, cells in the costal space, but this space does reach five cells wide in O. worleyae (see paratype 1, SR 16-006-001, Fig. 40 View FIGURE 40 ). The Okanagrion threadgillae ( Figs. 19 View FIGURE 19 , 20 View FIGURE 20 ) and O. lochmum ( Fig. 23 View FIGURE 23 ) holotypes have five cells wide in the RA–RP1 space, and while O. hobani specimens usually have a maximum of four cells wide in this space, it does reach this remarkable number in paratype 3 (SR 94-05-22, Fig. 27 View FIGURE 27 ), paratype 4 (SRUI 99-96-55, Fig. 28 View FIGURE 28 ), and paratype 6 (SR 00-04-05, Fig. 30 View FIGURE 30 ). Garrouste et al. (2017) noted that five cells width in these two spaces distal to the pterostigma was then only known in the Odonata in V. multicellulata .
The pterostigma of V. multicellulata is also distinct by its shape, the distal side is strongly curved and the proximal side is very long and oblique to C and RA, with two crossveins between the Costa and proximal side of the pterostigma, i.e., hyperstigmal crossveins. Hyperstigmal crossveins are uncommon in extant Zygoptera ( Garrouste et al. 2017). The pterostigmata of Okanagrion species are shaped like this, and the majority of specimens of O. hobani , and one each of O. worleyae and O. beardi have a hyperstigmal crossvein, and the holotype hind wing of O. threadgillae bears two ( Fig. 20B View FIGURE 20 ). The pterostigma of Valerea is about three times longer than wide, within the lower range of Okanagrion . The vein fragment of V. multicellulata that we interpret as CuA is linear at least well proximal to the origin of IR1, as in Okanagrion .
The proximal portion of IR 2 in V. multicellulata is not preserved from immediately distal to the level of the nodus. Unlike the condition in Okanagrion species where IR2 originates at this level either closer to RP1-2 or on RP1-2, the proximal-most preserved portion in V. multicellulata is relatively far from RP1-2 and closer to RP3-4, indicating that it likely originates well proximal to the nodus, or if at that level, then on or near RP3-4.
The Thanetian of Menat and the Ypresian Okanagan Highlands are probably about three to six million years apart, and would have been connected by continuous land at that time, with mild climates reaching high latitudes facilitating intercontinental dispersal (reviewed: Archibald & Makarkin 2006; Archibald et al. 2011b).
The specimen of undetermined genus and species from the Thanetian or Ypresian Chickaloon Formation of Alaska ( Garrouste & Nel 2019) appears quite close to Okanagrion with its dense venation, including an expanded CuA–A space four cells wide at its widest, between the levels of origins of RS 2 and IR1. In its diagnosis, above, we cite the MA beginning to zigzag in the distal-most portion of preserved in the Chickaloon specimen as distinguishing it from Okanagrion (where this is linear), although this is not a great difference, and they might be congeners or closely related genera. It is clearly not a species of Valerea as, e. g., the crossvenation is much more dense and the origins of IR1 and RP2 are nearer to the nodus in that genus.
Species richness. It is not surprising that there is high species richness of Okanagrion at two localities: eight in total, with four at McAbee and five at Republic (one of these shared between them). High alpha diversity was established at McAbee by Archibald et al. (2010), who found insect and woody dicotyledon species richness there to be equivalent to that of a modern lowland tropical rainforest in Costa Rica. That this richness is not an artefact of splitting was confirmed by the independent assessments of experts in individual insect orders (see Archibald et al. 2010). Such high, tropical levels of alpha diversity for woody dicotyledons have also been established at Republic ( Wilf et al. 2003).
This is also consistent with the findings of Archibald et al. (2013) that insect beta diversity is high between site pairs in the Okanagan Highlands. They extended Janzen’s hypothesis “Mountain passes are higher in the tropics” to mid-latitude mountainous regions of the Eocene by their “greenhouse world” extra-tropical climatic equability. This diversity pattern is then expected with climate-driven high alpha diversity, and climate and topography-driven high beta diversity. Other genera described here are less speciose.
Wing colour patterning. There is sexual dimorphism in wing colouration in a variety of extant Zygoptera and this may also be true in Cephalozygoptera . Further, there may be differences in patterning between the hind wings and forewings. In some Zygoptera species, there is also colour polymorphism in a single sex (e.g., clear-winged and pattern-winged males of Paraphlebia (Thaumatoneuridae) , but this is uncommon.
There are 17 specimens of Okanagrion hobani illustrated and discussed here, one of which we know is a male. While it seems unlikely that all 17 would be males, still, this is possible. Okanagrion species concepts are defined by other wing traits, with wing colouration as supplementary. Taking a conservative stance, we identify wing colouration as male in the O. hobani diagnosis, with female colouration unknown, and in other species specify that the sex is unknown. Colouration is consistent between wings identified as fore- and hind wings, and in the articulated fore- and hind wings of O. hobani paratype 7, F-1044.
Forewings and hind wings. Although forewings are close in shape to hind wings in Cephalozygoptera , Anisozygoptera and Zygoptera , they may in cases be somewhat different (e.g., slightly in Paraphlebia zoe Selys in Hagen see Figs. 470 and 436 and 437 of Garrison et al. 2010, more so in Thaumatoneura inopinata McLachlan , their Figs. 470 and 471). We base our assignment of fore- and hind wings on those of the holotype (and only specimen) of O. threadgillae , where two wings that conform to the differences seen in some modern species are in close association: the wing that we presume to be a forewing is slightly wider at the base, with the posterior margin slightly convex, widened at the level of the distal portion of the quadrangle ( Fig. 35 View FIGURE 35 , red arrows), and the anterior margin is slightly curved between the nodus and the pterostigma. In the presumptive hind wing, the proximal posterior margin is smoothly curved through the level of the quadrangle ( Fig. 35 View FIGURE 35 , blue arrow), the anterior margin is rather straight from the nodus to the pterostigma, and the posterior margin is more deeply curved. We compare this difference to that of the fore- and hind wings of Thaumatoneura species, although not as pronounced as in some of those. Unfortunately, in the male specimen F-1044 of O. hobani with all four wings preserved, these are somewhat damaged, precluding meaningful comparison of details of shape. Rather complete and undamaged wings of Okanagrion species appear to vary in the same manner as do those that we presume to be the fore- and hind wing of O. threadgillae , and so we assume these identities where possible.



















No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.