
Oryzomys bauri Allen, 1892
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4144.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:8CC5C9D3-6575-433D-B6B5-CD2B1CE6B80A |
|
DOI |
https://doi.org/10.5281/zenodo.5624965 |
|
persistent identifier |
https://treatment.plazi.org/id/03B487F6-FF87-FFC0-FF56-FA06FD6DFD38 |
|
treatment provided by |
Plazi |
|
scientific name |
Oryzomys bauri Allen, 1892 |
| status |
|
Type material. The type of Oryzomys bauri , described by Allen in 1892, is an adult male, part of a series composed by additional four specimens in alcohol: “I male two-thirds grown, I male half grown, I female adult and I female about half grown”, all captured in Barrington Island. Allen (1892) also mentioned the existence of three skins preserved in antiseptic solution. These specimens were sent to J. A. Allen by Dr. G. Baur of Clark University , Worcester , Massachusetts, and were obtained during the Salisbury Expedition in the Galapagos Archipelago ( Allen 1892). Those specimens were probably loaned to the AMNH through J. A. Allen, however they are not housed at the AMNH, neither in the regular nor in the type collections. As a matter of fact, on the original description there is no information on where this material may have been deposited, or any mention to voucher field numbers or collection numbers. We contacted Clark University (Jamie Fitzgerald and Paula Kupstas in litt.), and were informed that most rodents deposited there lack labels and that most of their identifying information has been lost over the last decades. Therefore, we believe that the designation of a neotype is important to define the nominal taxon objectively. We support this claim, since we consider that the type is lost, and we believe that this attends the qualifying conditions established by the ICZN (1999) article 75.3.4. Moreover, we selected as a neotype, a specimen from the same type locality (article 75.3.6), that is housed at the Museum of Vertebrate Zoology, a traditional institution that houses several type specimens (article 75.3.7), attending ICZN (1999) requirements.
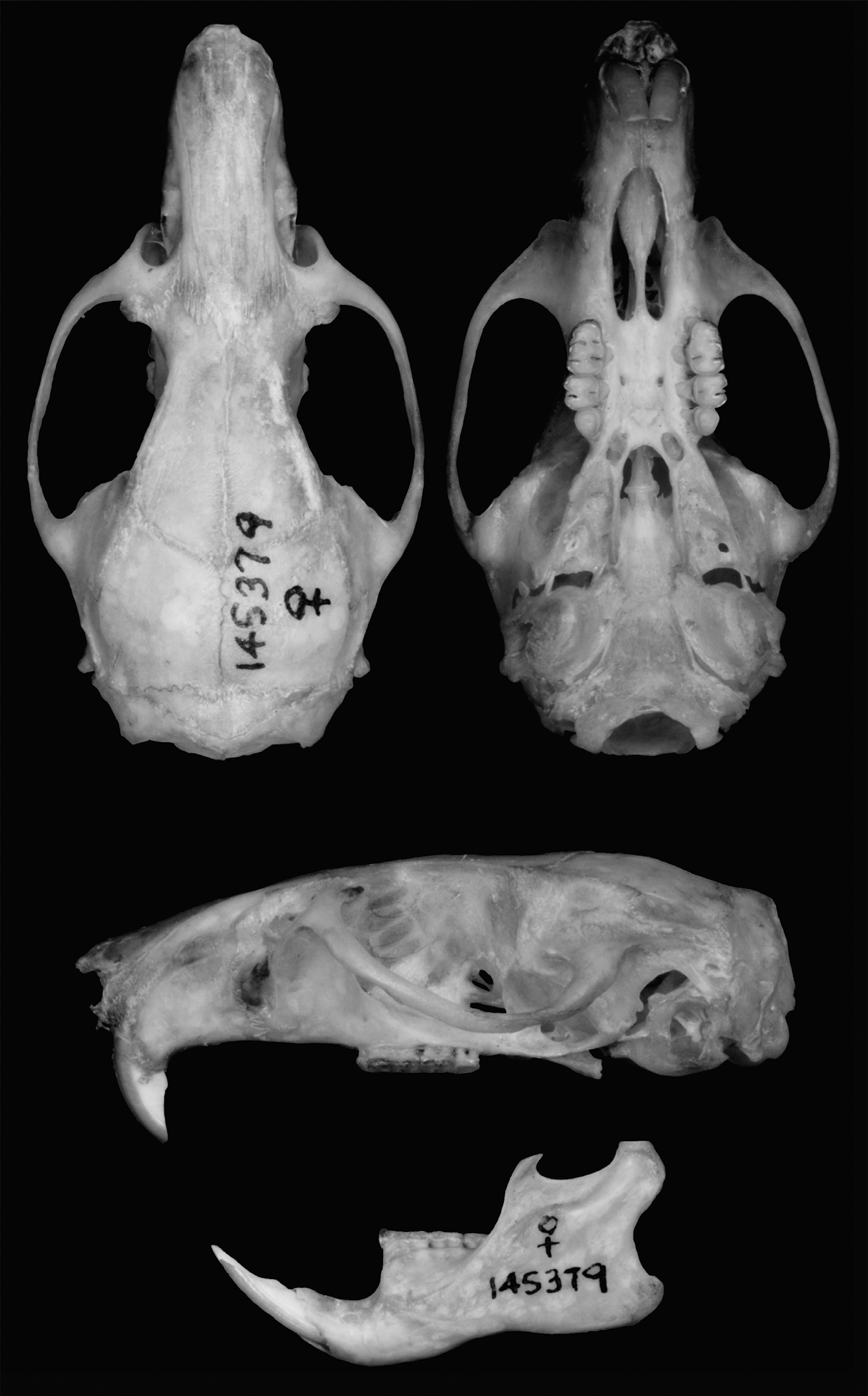
NEOTYPE: Adult female, deposited at the Museum of Vertebrate Zoology, under number MVZ 145379, collected by J. L. Patton, in 21.I.1974, number JLP 3951. It is preserved in skin, skull and tissue in ethanol; skin and skull are in good condition ( Fig. 4 View FIGURE 4 ). Other ten specimens are also part of the series, all deposited in the Museum of Vertebrate Zoology: five females, MVZ145372, MVZ145373, MVZ145376, MVZ145377, MVZ145379, and five males, MVZ145374, MVZ145375, MVZ145378, MVZ145380, MVZ145381. We chose this specimen since it is well preserved and harbors important information associated, such as skin, skull and tissue preserved on alcohol.
Type condition. Specimen preserved in skin and skull; both are in excellent condition, without any broken part.
Type locality. According to the original label, the neotype is (as the original missing specimens) from “ Barrington Cove , Isla Barrington ”. Here , we propose the following arrangement: “ Barrington Cove, Barrington Island [= Isla Santa Fe], Galapagos Archipelago, Ecuador ; geographical coordinates 00°49’S, 90°04’W ”. The type locality is represented in figure 3. The specimens employed by Allen (1892) in the description of this species were originally collected in “Barrington Island ” GoogleMaps .
Original description. The original description of the type designated by J. A. Allen was published in the Bulletin American Museum of Natural History, vol. 4, Article 4, 1892 (48–49), and is presented below:
“Evidently allied to Oryzomys galapagoensis (Waterh.) , but differing from it in proportions and coloration.
Pelage full and long. General color above dusky grayish brown, faintly varied with pale yellowish brown; below the pelage is white at the surface, passing into plumbeous. Muzzle lighter and more grayish, tinged strongly with yellowish brown on the sides of the muzzle and faintly so on the chin. Ears large, obtusely rounded, almost naked within, well haired externally on the anterior third, and sparsely haired over the rest of the outer surface. Feet above thinly haired, white faintly tinged with yellowish; soles naked, 6-tuberculate; posteriorly smooth and pale horn color, anteriorly granulated and yellowish, slightly varied with gray. Tail indistinctly bicolor, nearly naked, the annulations showing distinctly through the very short hairs; above dusky brown, below ashy brown.
Measurements —Male Adult: Total length, 280 mm; head and body, 132; tail, 147; hind foot, 32.5; fore foot, 18; ear, 18. Female Adult: Total length, 269; head and body, 124; tail, 145; hind foot, 31; fore foot, 16.5; ear, 17.3. Young Male: Total length, 248; head and body, 108; tail, 138; hind foot, 16.5; fore foot, 14.5; ear, 16. 5.
Skull —Total length, 37.4; basal length (condyles to incisors), 27.5; greatest zygomatic breadth, 18.3; least interorbital breadth, 5.6; length of nasals, 13.5; from posterior border of palate to incisors, 14.7; length of lower jaw (tip of incisors to condyle), 22.8; height at condyle, 9.9.
Based on 5 specimens in alcohol, namely, I male adult, I male two-thirds grown, I male half grown, I female adult, and I female about half grown, taken on Barrington Island, July 9 and I0, I89 I. There are also three inverted skins in antiseptic solution, taken at the same place and time. The adult male may be considered as the type ”
Dimensions of the neotype. External: Total length: 308 mm; length of tail: 160 mm; hindfoot: 32 mm; length of ear: 23 mm; weight: 94g; Craniodental: measurements of the neotype are available in table 2.
Morphological description of the neotype. Dorsal pelage very long, lax, soft and woolly; yellowish wool and cover hairs and brown guard hairs; dorsal coloration yellowish cream, weakly and grossly grizzled with dark brown resulting in brownish coloration; venter predominantly gray colored; tail greater than length of body, densely covered by long hairs, weakly bicolored, with large scales, about 16 scales/cm; pes densely covered by long hairs, with ungual tufts dense and longer than claws, all pure white; interdigital and plantar pads fleshy, hypothenar and thenar pads closer to the calcaneus and densely covered with squamae; pinnae externally covered with long hairs.
Skull moderately robust, with long and narrow rostrum (approximately 39% of skull length [LN/ONL ratio]); zygomatic plate moderately projected anteriorly, not reaching beyond nasolacrimal capsule; rostral fossa moderately deep; moderately projected plate and deep fossa, configuring a moderately deep and wide zygomatic notch; interorbital region convergent anteriorly, with supraorbital margins sharp and acute, forming small dorsolateral crests; zygomatic arches weakly divergent posteriorly, wider near the squamosal root; braincase small and delicate, elongated; braincase with temporal margins slightly squared, without crests; interparietal short and wide; fronto-squamosal suture contiguous with fronto-parietal suture; alisphenoid strut absent; anastomotic channel present, configuring pattern 3 of carotid circulation ( Voss 1988); parietals expanding over surface side of skull; postglenoid foramen relatively large; tegmen tympani small, not overlapped to squamosal; incisive foramen long, wider medially, with anterior and posterior margins rounded, posterior margin reaching anterocone of M1; posterior margin of zygomatic plate situated anterior to the alveolus of M1; intermediate palate, mesopterygoid fossa extends anteriorly between the maxillary bones but not between molar; palate with deep posterolateral palatal pits recessed in deep and large palatine depressions; palatal excrescencies weakly developed; anterior border of mesopterygoid fossa rounded, roof of mesopterygoid fossa perforated by very large vacuities, occupying the presphenoid and basisphenoid; auditory bulla, with short and wide stapedial process.
Upper incisors opisthodont; upper and lower molars with labial and lingual cusps arranged in opposite pairs; dental series with excessive wear, hindering the full dental description (we provide a description of the molars of the neotype series on the item “Comparisons among type specimens”). M1 with anteroloph almost entirely coalesced to anterocone; mesoloph partially fused to paracone (labial portion not fused); M2 with mesoloph partially fused to paracone (labial portion not fused); M3 small and subtriangular.
Observation. In the original description, Allen (1892) pointed out the close relationship between O. bauri and O. galapagoensis , being the first species later synonymized with Mus galapagoensis by Cabrera (1961). Nevertheless, Allen justified the validity of a new species by the fact that, even though individuals of O. bauri have similar size as those of O. galapagoensis , O. bauri representatives exhibit larger ears and tails longer than head and body, and, furthermore, coloration much less grizzled with yellow.
On the geographical distribution and habitat of the species, Dr. Baur (cited by Allen 1892: 50), stated that:
“On Barrington Island the small Rodent was pretty common; it was found between the bushes near the shore, and also high up between grass and the lava rocks.”
| AMNH |
American Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.