
Roboastra gracilis, (BERGH, 1877)
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00167.x |
|
DOI |
https://doi.org/10.5281/zenodo.5113670 |
|
persistent identifier |
https://treatment.plazi.org/id/03B5622C-FF92-1202-FC3E-8145FD32F9D6 |
|
treatment provided by |
Carolina |
|
scientific name |
Roboastra gracilis |
| status |
|
ROBOASTRA GRACILIS ( BERGH, 1877) View in CoL
( FIGS 1 View Figure 1 , 2 View Figure 2 , 3A View Figure 3 , 4A View Figure 4 )
Nembrotha gracilis Bergh, 1877: 458 View in CoL , pl. 56, figs 11- 17.
Nembrotha rubropapulosa Bergh, 1905: 198 View in CoL , pl. 5, fig. 14, pl. 18, figs 19-22.
Material examined: Cebu Island : White Beach ( Moalboal area ), Philippines, April 1996, 1 specimen, 0– 21 m depth, collected by T. M. Gosliner ( CASIZ: 106591). Marshall Islands: Enewetak North, Pacific Ocean, February 1982, 1 specimen, 1 m depth, collected by S. Johnson under dead coral ( CASIZ: 120670) . Koror Municipality : Ngederrak Lagoon, Palau, September 1996, 2 specimens, collected by Clay Carlson ( CASIZ: 109707). North coast, near Madang, Barracuda Point , Papua New Guinea, November 1990, 2 specimens, 6 m depth, collected by T. M. Gosliner ( CASIZ: 075839). North coast, near Madang, Christiansen Research Institute’s Clam Tanks , Papua New Guinea, November 1991, 1 specimen. collected by T. M. Gosliner ( CASIZ: 086900). North coast, near Madang, outer barrier reef between Pig Island and Rasch Pass, Papua New Guinea, June 1992, 1 specimen, collected by T. M. Gosliner ( CASIZ: 086496). Luzon , Batangas Province, Maricaban Island , Philippines, 1 specimen, collected by M. Miller ( CASIZ: 103732). Molokini Island , Hawaii , April 1990, 1 specimen, 6 m depth, collected by P. Fiene under rubble. Identified as ‘ Roboastra n. sp., aff. gracilis’ ( CASIZ: 087131). Maui , Hawaii , April 1990, 1 specimen, 10 m depth, collected by P. Fiene. Identified as ‘ Roboastra sp. like the one in Scott Johnson’s book’ ( CASIZ: 088547). Oahu Island R. cf. gracilis and Makua , Hawaii , June 1980, 1 specimen, 5 m depth, collected by S. Johnson and identified as ‘-light background’ ( CASIZ: 116819). Kimberley , Ashmore Reef, Western Australia, July 1994, 1 specimen, 5–15 m depth, collected by C.W. Bryce ( WAM: S23267 View Materials ) . North of Cartier Island, Western Australia, September 1988, 1 specimen, collected by C.W. Bryce & F.E. Wells ( WAM: S23236 View Materials ) . Off Fremantle , Hall Bank, Western Australia, December 1981, 1 specimen, collected by S.M. Slack-Smith & C.M. Bryce ( WAM: S23233 View Materials ) .
Distribution: Tropical Indo-Pacific: South Africa, Red Sea ( Saudi Arabia), Malaysia, Indonesia ( SE Sulawesi, Bali), Palau, Philippines (the type locality), Papua New Guinea, Solomon Is., Japan, Australia, Western Australia, New South Wales, N. Queensland, Pacific Ocean (Marshall Is.), Fiji and many other records ( Baba, 1936; Burn, 1967; Hamatani & Baba, 1976; Baba, 1977; Coleman, 1989, 2001; Ono, 1999, 2004; Suzuki, 2000; Takamasa, 2003; Nakano, 2004).
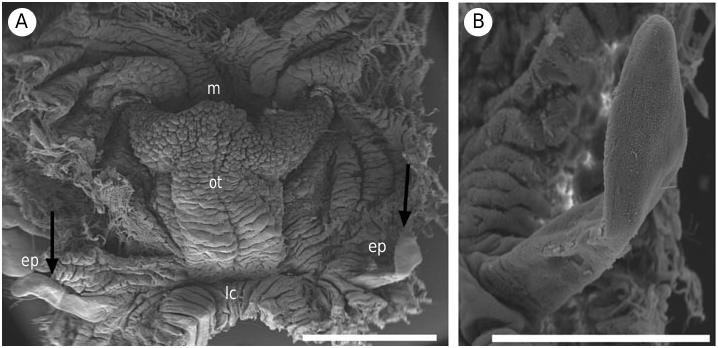
External morphology: Small, measuring 10–30 mm in length. Body elongate and limaciform with a long and pointed posterior end of the foot. The foot is linear and blue-grey. The ground colour of the body is blueblack with yellow-orange longitudinal lines, which can be continuous or discontinuous ( Fig. 1A, B View Figure 1 ). If the lines are discontinuous they give the appearance of yellow-orange spots. The longitudinal lines or spots are arranged in the following pattern: 2 stripes in the median part of the back between the head and the front of the gills, 2 in the lateral parts of the back between the rhinophores and the rear of the gills, 1 around the margin of the mantle and 3–4 on the sides of the body. Usually the yellow lines are narrower than the blue-black background. All these lines consist of a series of glandular pits, which give them a corrugated aspect ( Fig. 1C View Figure 1 ). The oral tentacles are smooth, very well developed and dorsolaterally grooved. The perfoliate rhinophores are very large, with 15–25 packed lamellae. There are from three to six very large and nonretractile unipinnate gills. The gills form a semicircle surrounding the anal papilla, which is not elevated. These proportionally large rhinophores and gills are a very characteristic feature of this species. The genital pore opens on the right side, slightly closer to the rhinophores than to the gills. The oral tentacles, the rhinophores and the gills can be deep blue, blue-grey or purple depending on the specimens.
Internal morphology: The anterior digestive tract begins with a long and muscular oral tube that continues into the buccal mass. At the junction of the oral tube and the buccal mass a pair of thick elongated pouches opens into the digestive system. These pouches are arranged similarly in all Roboastra spp. ( Fig. 10A View Figure 10 ). There are two small, short salivary glands on the buccal bulb, flanking the oesophagus. The buccal mass is shorter in length than the oral tube. The labial cuticle is smooth and very weak. The radular formula of the two 15-mm-long specimens (preserved) is 30 ¥ 3–2.1.1.1.2–3; that of the 4-mm and 6-mm-long specimens (preserved) is 25 ¥ 2.1.1.1.2. The rachidian tooth is rectangular, with a slightly arched upper edge and three denticles. These denticles, while are usually the same size, can be highly asymmetrical ( Fig. 2A, B View Figure 2 ). The inner lateral tooth has typically a strongly curved ga bifid inner cusp. The upper denticle of this inner cusp is strong and little developed; the outer cusp is slender and well developed ( Fig. 2C View Figure 2 ). The proximal outer lateral teeth become smaller near the margin. The inner outer lateral teeth are longer and larger than the remaining outer lateral teeth ( Fig. 2A, B View Figure 2 ).
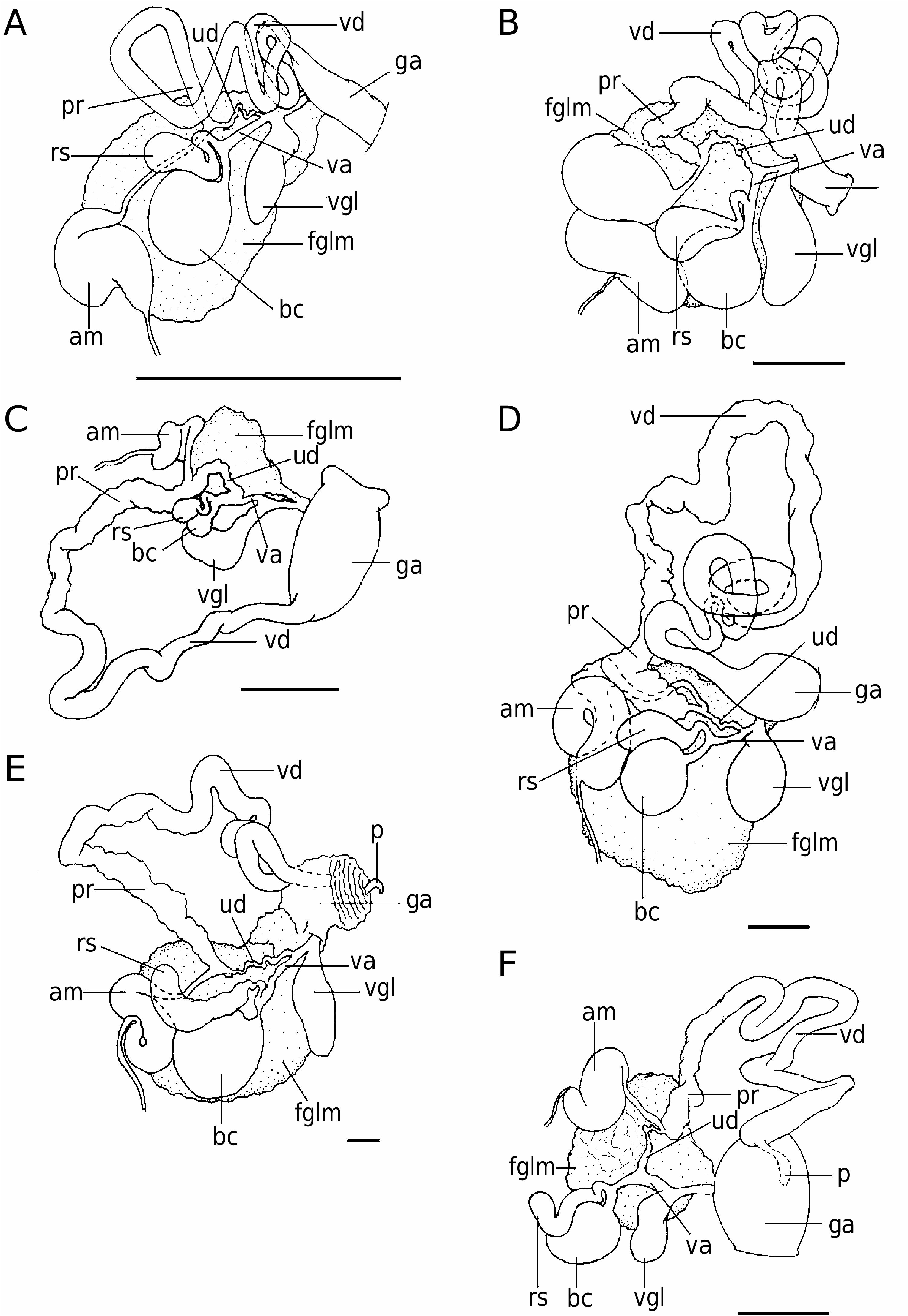
The reproductive system is triaulic ( Fig. 3A View Figure 3 ). The genital mass fills the whole of the second quarter of the body cavity. The hermaphroditic duct widens into a large S-shaped ampulla. The ampulla narrows into a long thin postampullary duct that connects with the oviduct and the prostate. The short oviduct enters the massive female gland mass. The deferent duct lacks a morphologically well-differentiated prostate but it has soft glandular walls that skirt around the seminal receptacle. The distal muscular vas deferens is much narrower and shorter than the prostatic part. It terminates in a dilated penial section. The penis is armed with at least three different kinds of hooked, chitinous spines arranged in helicoidal rows ( Fig. 4A View Figure 4 ). The vaginal duct is long, normally straight and connects to the large, round bursa copulatrix. The seminal receptacle, smaller and more elongate than the bursa copulatrix, has a short duct that connects to the vagina near the bursa after making two loops. A slender uterine duct leaves the vagina and joins the oviduct. The vagina shares a common aperture within the genital atrium with the vaginal gland, which is very well developed, elongate, flattened, with muscular walls.
-
Remarks: Bergh (1877) described Roboastra gracilis (as Nembrotha gracilis ) based on specimens collected from the Philippines. His types were two specimens preserved in alcohol; there are neither notes on, nor sketches of, the living animals. Baba (1949) depicted another specimen from Japan and said that N. gracilis may have either two or three marginal plates. Burn (1967) discovered two new specimens from northern New South Wales ( Australia) and redescribed the species under the name Roboastra gracilis justified by radular and genital differences. He included a drawing of the living animal made from a colour transparency and drawings of the half row of radula and the reproductive system.
Hamatani & Baba (1976) made a taxonomic comparison between R. gracilis and R. luteolineata using 16 specimens of the former, collected from Japan. In these notes they included a drawing of one specimen and two drawings of the left half row of the radula. No comment was made about the reproductive system. Later Baba (1977) described R. gracilis as being collected from Seto, Japan. He described in further detail both the colour and radula of this small animal; however, again there was no description of the reproductive system.
Although Burn’s and Baba’s descriptions of R. gracilis are not very detailed, they are consistent with our specimens. This species has very characteristic features: extremely long rhinophores and gills (the latter simply pinnate), which are unique within the genus, while the body surface is covered with glandular pits. In recent years, many pictures have appeared in different field books and web pages devoted to sea slugs and marine invertebrates. It has been reported that when the animals are disturbed, the yellow-orange spots along the body open, releasing an acidic yellow fluid ( Fig. 1C View Figure 1 ). Once the chemical has been released, they close the spots again. Presumably, this is a defence mechanism but it appears to be quite noxious to other sea slugs ( Warren, 2000). At the moment there are no feeding records for this species. However, as a typical Roboastra , it most likely feeds on other polycerids.
In our studies we found specimens in CASIZ that have different colours (but the same pattern of coloration) and other differences in their external anatomy ( Fig. 1D View Figure 1 ). They were collected from Hawaii and are 6 mm in length (preserved). Their ground colour is transparent white with lines or spots of brilliant yellow. The end of the oral tentacles, the tips of the rhinophores and gills and the end of the foot are purple. They have 3–4 simply pinnate gills, although these are shorter than in larger specimens from other areas within the Indo-Pacific. Initially, we thought that they might be a different species, but study of their internal anatomy revealed that they are R. gracilis . They are similar in colour to the specimens illustrated by Bertsch & Johnson (1982) and Suzuki (2000).
Roboastra rubropapulosa View in CoL was the second species of the genus described by Bergh (1905). It too has an Indo-Pacific distribution. In his report on the Siboga material, Bergh (1905: 198) described a brightly coloured papillate species ( Fig. 5A View Figure 5 ) as N. rubropapulosa View in CoL . This differed from his R. gracilis View in CoL only in that there was one elongate marginal plate on each side of the radular rows. Burn (1967) stated that the number of plates is not a characteristic which can be used to separate the species from its congeners because the division between the marginal plates can be very difficult to see. Thus N. rubropapulosa View in CoL cannot be effectively separated from R. gracilis View in CoL and is presently considered to be a synonym of the latter.
Nevertheless, photographs have appeared in a number of publications (e.g. Gosliner, Behrens & Willians, 1996) and in web pages (www.medslug.de; www.diveoz.au.com), where the animal is misidentified as R. rubropapulosa View in CoL ( Fig. 5B View Figure 5 ). Moreover, we have studied some of the specimens deposited at CASIZ (075236, 082920, 082922, 106531) and we have been able to confirm that they represent juvenile specimens of Nembrotha sp., perhaps N. kubaryana View in CoL , because they have a radula typical of this genus ( Fig. 5C View Figure 5 ).




| T |
Tavera, Department of Geology and Geophysics |
| R |
Departamento de Geologia, Universidad de Chile |
| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Roboastra gracilis
| Pola, M., Cervera, J. L & Gosliner, T. M. 2005 |
Nembrotha rubropapulosa
| Bergh LSR 1905: 198 |
Nembrotha gracilis
| Bergh LSR 1877: 458 |