
Hoplodoris, BERGH, 1880
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2002.00039.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B5879A-756F-6C78-90BE-F9CB2150AE42 |
|
treatment provided by |
Carolina |
|
scientific name |
Hoplodoris |
| status |
|
GENUS HOPLODORIS BERGH, 1880 View in CoL
Hoplodoris Bergh, 1880: 51 View in CoL . Type species: Hoplodoris desmoparypha Bergh, 1880 View in CoL , by monotypy.
Diagnosis
Dorsum covered with simple, large and rounded tubercles, stiffened by integumentary spicules. Head with two conical oral tentacles. Anterior border of the foot grooved and notched. Labial armature armed with jaw elements. Radula composed of simple, hamate teeth, occasionally denticulate. Reproductive system with a flattened, granular prostate, having two well differentiated regions. Penis armed with hooks. Vagina devoid of armature. One or two large and pedunculated accessory glands armed with copulatory spines.
Remarks
Bergh (1880) described the genus Hoplodoris based on Hoplodoris desmoparypha Bergh, 1880 , the type species by monotypy. The genus Hoplodoris is characterized by having the dorsum covered with long tubercles, the anterior border of the foot grooved and notched, presence of jaws, hamate radular teeth, large prostate, penis armed with hooks and presence of an accessory gland with a spine. The type species of Hoplodoris has not been collected since, and there is no information on its external morphology. Unfortunately, the type material of this species collected from Palau could not be located at ZMUC and is presumed lost. The information for this genus used in the phylogenetic analysis has been obtained from Hoplodoris novaezelandiae ( Bergh, 1904) .
Thompson (1975) regarded Carminodoris as a junior synonym of Hoplodoris based on his description of Hoplodoris nodulosa Angas, 1864 from Australia. However, the original description of Carminodoris ( Bergh, 1889) , based on Carminodoris mauritiana , states that this genus is characterized by having the anterior bor- der of the foot grooved and notched, the dorsum covered with small tubercles, presence of jaws, hamate lateral teeth, denticulate outermost lateral teeth, large prostate and penis armed with hooks. As mentioned below, it is probable that Carminodoris , which lacks accessory glands and has small dorsal tubercles, is a synonym of Discodoris , but this point needs confirmation.
Other genera having accessory glands with spines are Asteronotus Ehrenberg, 1831 ; Jorunna Bergh, 1876 and Paradoris Bergh, 1884 . According to the results of the phylogenetic analysis (see below), all these taxa belong to different clades, and it is very likely that they acquired the copulatory spines independently.
Several species have been added to the genus Hoplodoris since its original description. Burn (1969) transferred Doris nodulosa Angas, 1864 , Miller (1991) transferred Homoiodoris novaezelandiae Bergh, 1904 and Gosliner & Behrens (1998) described the new species Hoplodoris estrelyado Gosliner & Behrens, 1998 . Some Indo-Pacific species previously assigned to the genus Carminodoris Bergh, 1889 , should also probably be transferred to Hoplodoris (see Gosliner & Behrens, 1998). Gosliner & Behrens (1998) described some variation within Hoplodoris to accommodate species with one or two accessory glands armed or unarmed with spines. All the species included in Hoplodoris are characterized by having rounded dorsal tubercles, except for Hoplodoris desmoparypha , which has elongate tubercles.
HOPLODORIS NOVAEZELANDIAE ( BERGH, 1904) View in CoL
( FIGS 32 View Figure 32 , 33 View Figure 33 )
Homoiodoris novaezelandiae Bergh, 1904: 35–37 , pl. 3, figs 3–7.
Type material
SYNTYPES: Port Chalmers , New Zealand, date unknown, four specimens, 10–12 mm preserved length, leg. H. Suter ( ZMUC GAS-2105 ) .
External morphology
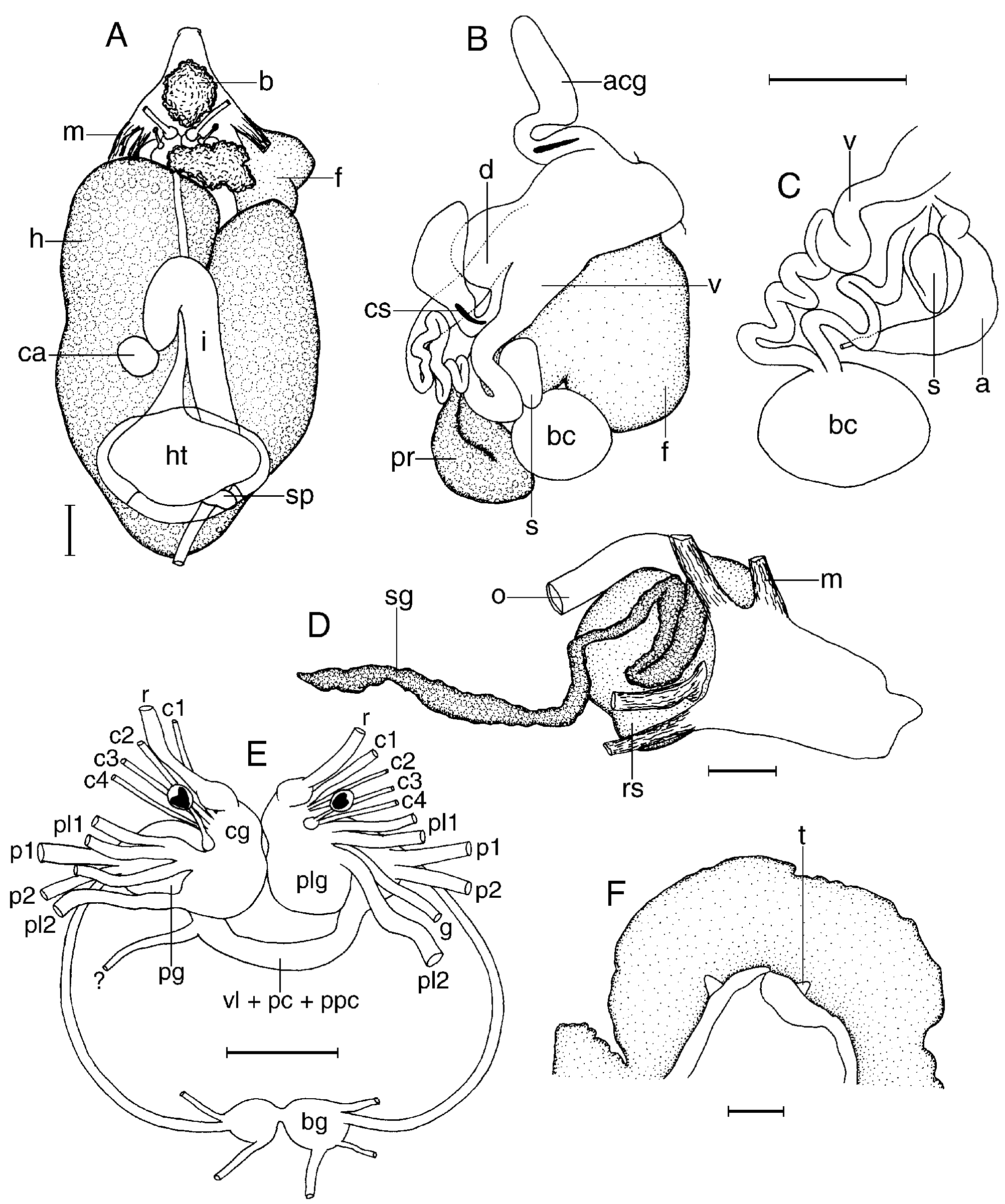
The animals here examined were preserved, so no information on the external coloration was available. The external morphology of this species has been described and illustrated by Miller (1991). In the examined specimens the entire dorsum is covered with large, rounded tubercles ( Fig. 32F View Figure 32 ). Some larger tubercles are randomly distributed among the others. The rhinophoral and branchial sheaths have papillae similar to those on the rest of the dorsum. There are 10 tripinnate branchial leaves. The anal papilla is situated in the centre of the branchial circle of leaves. The rhinophores are elongate, having 14 lamellae in a 12-mm preserved length specimen.
Ventrally the anterior border of the foot is grooved and notched ( Fig. 33F View Figure 33 ). The oral tentacles are conical.
Anatomy
The posterior end of the glandular portion of the oral tube has six strong retractor muscles ( Fig. 33D View Figure 33 ) which attach to the body wall. The oval, muscular buccal bulb has two large additional muscles attached; two long salivary glands connect with it at each side of the oesophageal junction. The buccal bulb is shorter than the glandular portion of the oral tube. The labial cuticle is armed with thin elements ( Fig. 32D View Figure 32 ). The radular formula is 40 ¥ 25.0. 25 in a 12-mm preserved length specimen. Rachidian teeth are absent. The innermost lateral teeth are hamate and have up to seven irregular denticles on the inner side of the cups ( Fig. 32A View Figure 32 ). The next lateral teeth are hamate and lack denticles ( Fig. 32B View Figure 32 ). The teeth from the middle portion of the half-row are larger than those closer to the medial portion of the radula. The outermost teeth are smaller and have irregular denticles ( Fig. 32C View Figure 32 ). The oesophagus is long and connects directly to the stomach.
The ampulla is short ( Fig. 33C View Figure 33 ). It branches into a short oviduct and the prostate. The oviduct enters the female gland mass near to its centre. The prostate is short and flattened ( Fig. 33B View Figure 33 ) and has two different portions that are clearly distinguishable in colour and texture. It connects with a long duct that expands into the large ejaculatory portion of the deferent duct. The penis is armed with a series of small hooks ( Fig. 32E View Figure 32 ). The muscular deferent duct opens into a common atrium with the vagina. There are two large and pedunculated accessory glands connected to the atrium, each one bearing a copulatory spine. At its proximal end the vagina joins the bursa copulatrix. From the bursa copulatrix leads another duct connecting to the uterine duct and the seminal receptacle. The bursa copulatrix is oval in shape, about five times as large as the elongate seminal receptacle.
In the central nervous system ( Fig. 33E View Figure 33 ) the cerebral and pleural ganglia are fused and distinct from the pedal ganglia. There are four cerebral nerves leading from each cerebral ganglion and two pleural nerves leading from each pleural ganglion. There is no separate abdominal ganglion on the right side of the visceral loop. The buccal ganglia are near to the rest of the central nervous system, joined to the cerebral ganglia by two relatively long nerves. Rhinophoral and optical ganglia are present. The pedal ganglia are clearly separated, having two nerves leading from each one. The pedal and parapedal commissures are enveloped together with the visceral loop.
The circulatory system ( Fig. 33A View Figure 33 ) consists of a large heart and two blood glands situated in front of and behind the central nervous system.
Remarks
Miller (1991) redescribed this species based on newly collected specimens from New Zealand. He examined all the New Zealand species of dorids known to exist and no other species come at all close to the specimens he re-described as Hoplodoris novaezelandiae . Even though he was sure that his specimens were correctly examined he found some differences with Bergh’s (1904) description. Re-examination of the type material of this species confirms that Miller (1991) identified his specimens correctly, and that the anatomy of the syntypes of Hoplodoris novaezelandiae examined here is identical to that of his specimens.

| ZMUC |
Zoological Museum, University of Copenhagen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Hoplodoris
| Valdés, Ángel 2002 |
Homoiodoris novaezelandiae
| Bergh R 1904: 37 |
Hoplodoris
| Bergh R 1880: 51 |