
Pseudocohnilembus persalinus Evans and Thompson, 1964
|
publication ID |
https://doi.org/10.1080/00222930701877540 |
|
persistent identifier |
https://treatment.plazi.org/id/03B587B3-FF88-C442-FE88-620AFBCBFE89 |
|
treatment provided by |
Felipe |
|
scientific name |
Pseudocohnilembus persalinus Evans and Thompson, 1964 |
| status |
|
Pseudocohnilembus persalinus Evans and Thompson, 1964 View in CoL
( Figures 3A View Figure 3 , 8J–N View Figure 8 )
Numerous cells were impregnated with both protargol and silver-nitrate methods. These isolates correspond perfectly to the redefinition by Song (2000) so that it is unnecessary to give a detailed redescription.
Description
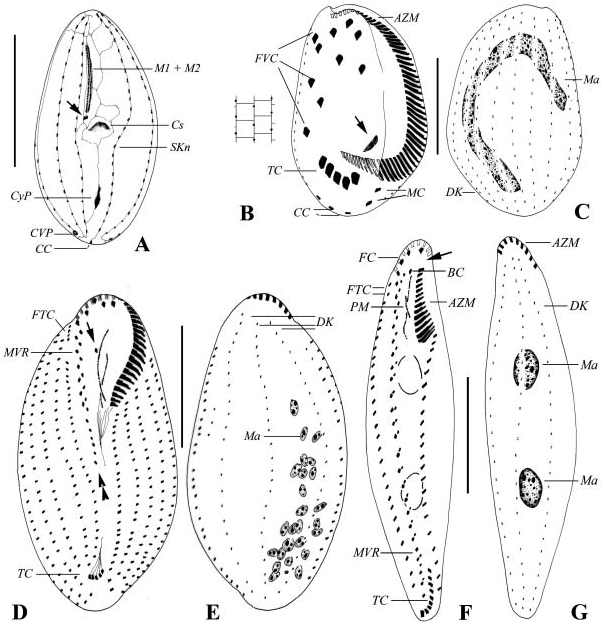
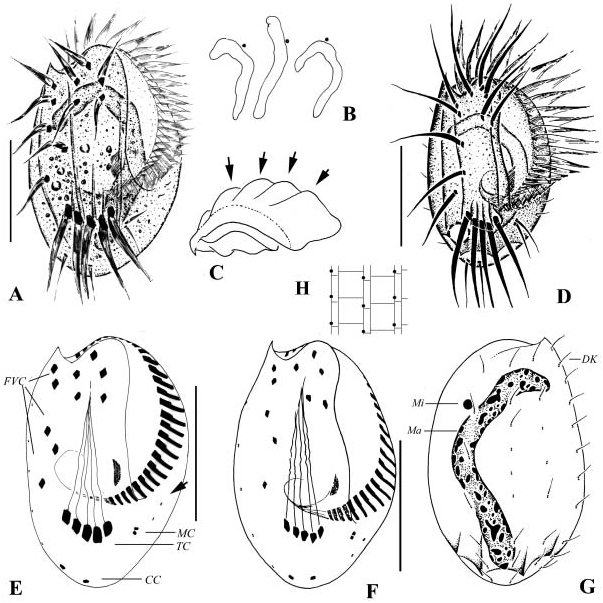
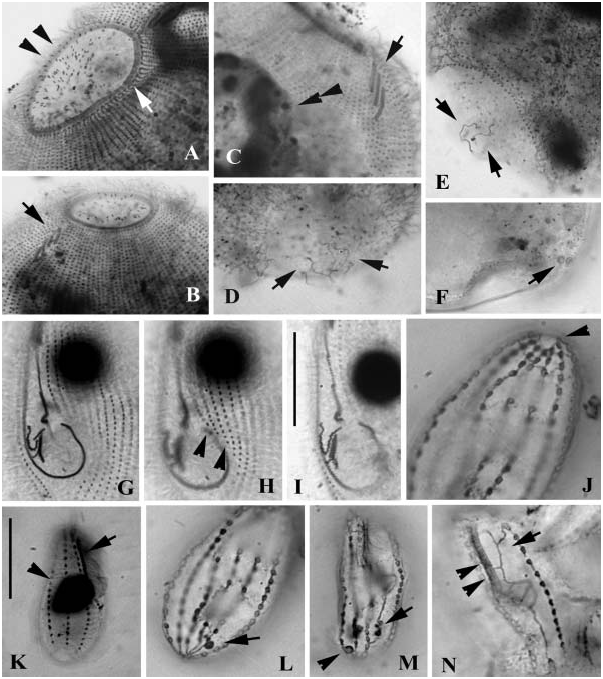
Cell size about 30–40×20–25 Mm after silver impregnation; body shape as shown in Figure 3A View Figure 3 , similar among individuals: anteriorly tapering, no distinct frontal plate. Area of buccal cavity wide, extending to about 1/2 of cell length. One large spherical macronucleus centrally located ( Figure 8K View Figure 8 , arrowhead), micronucleus not observed. Buccal apparatus genus-typical ( Figure 3A View Figure 3 ). Nine to 10 bipolar somatic kineties arranged sparsely, which are usually dikinetids in the anterior three-quarters of the body. About 20 basal bodies (or basal-body pairs) in each kinety. Silverline system as shown in Figures 3A View Figure 3 , 8J, N View Figure 8 , extrusomes not observed. Line along kinety No. n (first kinety to left of buccal field) continuing to caudal cilium complex, crossing through polar circle. Cytopyge (CyP) is thick argentophilic patch, located subcaudally ( Figures 3A View Figure 3 ; 8M View Figure 8 , arrow). Contractile vacuole pore (CVP) positioned invariably at end of 3rd somatic kinety ( Figures 8L View Figure 8 , arrow, in M, arrowhead).
Remarks
This organism differs from the congener Pseudocohnilembus hargisi in body shape (oval versus slender), lower number of somatic kineties (9–10 versus 13–14) and the position of CV pore (consistently at the posterior end of third kinety versus posterior end between fourth and fifth kineties) ( Gong et al. 2002; Song and Wilbert 2002b).
Pleuronema coronatum Kent, 1881
( Figures 8G–I View Figure 8 )
This species was also identified from protargol-impregnated specimens (with many individuals). All morphometric data correspond perfectly to the previous descriptions ( Dragesco 1968; Foissner et al. 1994; Song and Wilbert 2002a). Hence only some extra points will be supplied here.
Size rather variable, 60–90 Mm in length, yet body shape consistent as seen in silver-impregnated specimens. Thirty-four to 37 somatic kineties. Micronucleus always globular, similar in size, and centrally positioned. Oral apparatus typically as described previously ( Figures 8G, I View Figure 8 ; see also Song and Wilbert 2002a). Always four preoral kineties in the present sample ( Figure 8H View Figure 8 , arrowheads).
Order HETEROTRICHIDA Stein, 1859 Genus Bickella nov. gen.
Diagnosis for new genus
Benthic Folliculinidae with Folliculina -typical peristomial lobes and ciliary structure; without lorica, marine habitat.
Type species
Bickella antarctica nov. spec.
Dedication
We dedicate this new genus to Professor Dr. Hartmut Bick, who in the 1960s established the Ciliatology School of Bonn University, from which both present authors graduated, and who has greatly contributed to the ecology of ciliates.
Comparison and remarks
This new, monotypic genus differs from all other folliculinids in the absence of the lorica and the style of free-swimming behaviour. Although the type species (see below as well) was found in large abundance by the authors during the 2002–2004 surveys, we were not quite sure whether this organism really does not have the lorica. Subsequent examination of many samples confirmed that this organism is a true lorica-free form, and therefore should be considered an undescribed taxon. It is reasonable to believe as well that it could be closely-related to other lorica-possessing folliculinids; for instance, the genus Heterostentor is similar to the typical periphytic Stentor , which was also found in the Antarctic area by the present authors ( Song and Wilbert 2002a). The phenomenon of being ‘‘lorica-free’’ might represent an adaptation to the food-shortage biotope in Antarctic waters, i.e. organisms must more actively search for food in such an area.
Bickella antarctica nov. spec.
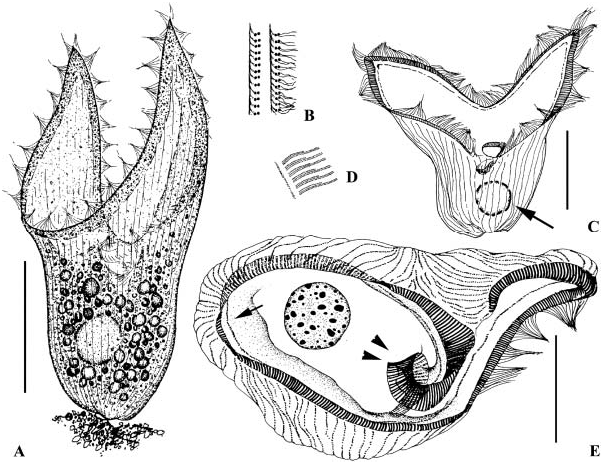
( Figures 4A–E View Figure 4 )
This species was initially described by Wilbert and Song (2005) as a unknown morphotype ( Folliculina ? sp., see Wilbert and Song 2005, Figure 6 View Figure 6 ). We re-collected
and re-checked this organism and, based on both isolates, we establish a new species here.
Diagnosis
Bickella in vivo about 250 Mm long; two prominent peristomial lobes of approximately equal size; adoral zone of membranelles containing about 850–1250 membranelles, ca. 50–110 somatic kineties in mid-body; single globular macronucleus about 30 Mm across.
Type locality
Benthos samples from rockpools on the sea coast near Bellinghausen Station (62 ° 139S, 58 ° 589W) and in the littoral zone of Potter Cove (62 ° 149S, 58 ° 40W 9), King George Island .
Description
Body overall (including peristomial lobes) about 150–400Mm long. Body shape and size rather consistent in all populations observed, cells not very sensitive to disturbance or contractile; two ear-shaped wings wide and prominent, generally transparent except in the border area, where membranelles are arranged. Cytoplasm colourless, usually dark grey due to numerous food granules, especially under lower magnification. Macronucleus globoid, about 30 Mm across with many large nucleoli; micronuclei not detected. Two forms of locomotion observed: swimming slowly in the water without features, or using the caudal end to attach at the bottom ( Wilbert and Song, 2005).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |