
Holosticha diademata (Rees, 1884)
|
publication ID |
https://doi.org/10.1080/00222930701877540 |
|
persistent identifier |
https://treatment.plazi.org/id/03B587B3-FF8F-C45B-FE78-6289FB76FC78 |
|
treatment provided by |
Felipe |
|
scientific name |
Holosticha diademata (Rees, 1884) |
| status |
|
Holosticha diademata (Rees, 1884) View in CoL
( Figures 12G, H, J View Figure 12 ; Table 3)
This species has been repeatedly found in the Antarctic area during recent years ( Petz et al. 1995; Song and Wilbert 2002a). Based on the impregnated specimens, a brief mention of some morphological variations is presented here.
Description
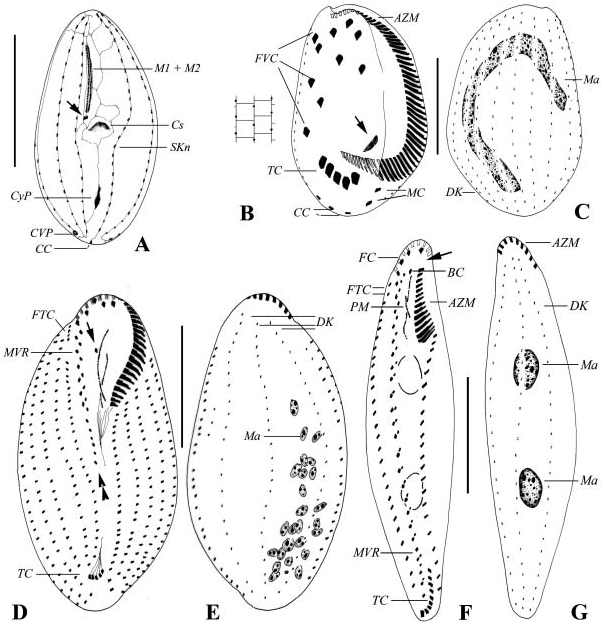
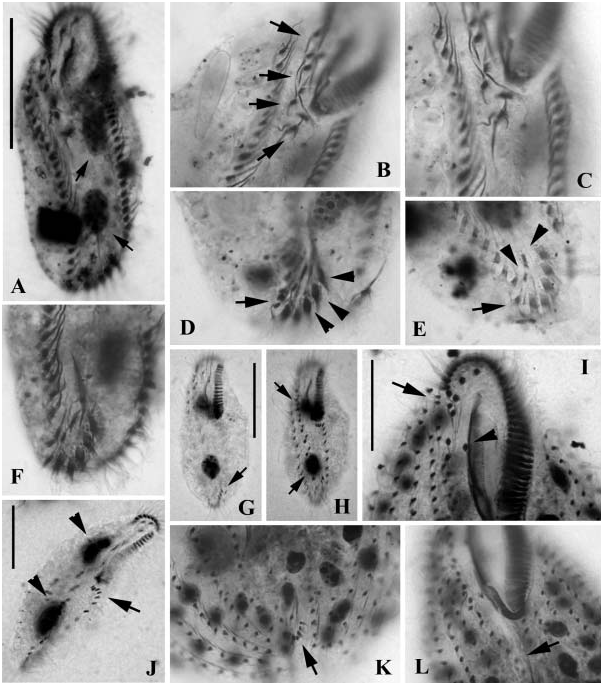
Body shape in the present sample is generally of two types: slender and elongate form (about 80×30 Mm in size), or oval form (about 60×30 Mm in size), both of which have identical infraciliature pattern ( Figures 12G, H, J View Figure 12 ). Basic ciliature features correspond unambiguously to those described before ( Petz et al. 1995; Song and Wilbert 2002a). What should be mentioned is that the bases of cirri on the ventral side in the current isolate differ slightly from the 2000 population ( Song and Wilbert 2002a), being comparatively finer or weaker.
Remarks
With the characteristic infraciliature and the nuclear apparatus, this species is easily identified ( Hu and Song 1999). Recently, Song and Wilbert (2002a) suggested that the form named Holosticha pullaster by Petz et al. (1995) could be conspecific with Holosticha diademata for two main reasons: (1) the former is a freshwater form as revised by Foissner et al. (1991); and (2) all five morphotypes found in the Antarctic area exhibit rather variable statistical data, which indicates that specimens of this contractile vacuole. Data after protargol impregnated specimens.
species could be highly population- or even individual-dependent in such extreme conditions ( Table 3). We maintain this opinion and await further evidence.
Holosticha apodiademata nov. spec.
( Figures 3F, G View Figure 3 ; Table 3)
Diagnosis
Band-like, marine Holosticha about 200–300 Mm long in vivo; adoral zone conspicuously bipartite with c. 8 anterior and 15 posterior membranelles; approximately 25 pairs of cirri in midventral rows which terminate at the transverse cirri; about 35 marginal cirri on both sides; four frontal, one buccal, two frontoterminal and 8–11 transverse cirri; consistently three dorsal kineties; two macronuclear nodules widely separated with 2–3 micronuclei. Cortical granules absent.
Type locality
In benthos samples from rockpools and the littoral zone on the sea coast near Bellinghausen Station, King George Island (62 ° 139S, 58 ° 589W).
Description
Form band-shaped, contractile and could be up to over 300 Mm long, though we failed to obtain detailed live information. Buccal field narrow and comparatively short, about 1/5–1/4 of cell length in vivo ( Figure 3F View Figure 3 ). Contractile vacuole not observed. Consistently two oval macronuclear nodules in widely separated positions, 2–3 spherical micronuclei recognized in protargol-impregnated specimens.
Infraciliature as shown in Figures 3F, G View Figure 3 . Buccal field narrow. Adoral zone conspicuously bipartite, with over 10-Mm-wide gap ( Figure 3F View Figure 3 , arrow) and considerable displacement, i.e. the posterior portion shifted to right of buccal field and not in line with the anterior portion ( Figure 3F View Figure 3 ). Membranelles in typical diademata -pattern; that is, bases in proximal portion become conspicuously longer posteriad. Paroral region (PM, Figure 3F View Figure 3 ) very short compared with the long endoral. One buccal cirrus (BC) at anterior level of endoral.
About four frontal cirri (FC), with anterior three conspicuously enlarged ( Figure 3F View Figure 3 ). Two frontoterminal cirri (FTC) near anterior end of right marginal row and hence often difficult to recognize. Midventral rows (MVR) prominent with about 25 pairs of cirri, arranged in typical zig-zag pattern, terminating completely at rear end of cell. Transverse cirri (TC) slightly enlarged and arranged in J-shape ( Figure 3F View Figure 3 ). Marginal rows not confluent posteriorly. Anterior end of left marginal row not (!) transversely arranged. Dorsal kineties (DK) consistently three, densely ciliated (ca 4 Mm); without caudal cirri ( Figure 3G View Figure 3 ).
Remarks
Due to the combination of long band-shaped body, two macronuclei, the large size and the ciliary pattern on both sides, especially the bipartite adoral zone of membranelles, this species is clearly recognizable from the other four related known congeners (see Table 3).
Metaurostylopsis marina ( Kahl, 1932) View in CoL
( Figures 3D, E View Figure 3 ; 12I, K, L View Figure 12 )
The Antarctic isolation matches the original and the redescription very well ( Kahl 1932; Song et al. 2001). We thus supply here only some new information derived from the impregnated specimens.
Description
Body size about 100–160×60–90 Mm according to fixed materials. Generally six left and six right marginal rows on average; adoral zone about one-third of body length with ca. 30 membranelles; about seven pairs of cirri in midventral rows, single short ventral row extending about halfway along the cell with ca. six cirri ( Figure 3D View Figure 3 , double-arrowheads); 2–4 frontoterminal (FC), 1 buccal ( Figure 3D View Figure 3 , arrow), about four frontal and five transverse cirri (TC, Figures 3D View Figure 3 , 12K View Figure 12 ). Consistently three dorsal kineties (DK, Figure 3E View Figure 3 ).
Remarks
Considering the infraciliature, this species is similar to the congener, Metaurostylopsis rubra , which is a reddish, larger and slender form also found in the Antarctic area ( Song and Wilbert 2002a). Apart from the difference in life, it can be recognized by the lower number of frontoterminal cirri (2–4 versus 5–8) and adoral membranelles (,30 versus c. 40), as well as other statistical data (see also Song and Wilbert 2002a).
Hemigastrostyla szaboi Wilbert and Song, 2005
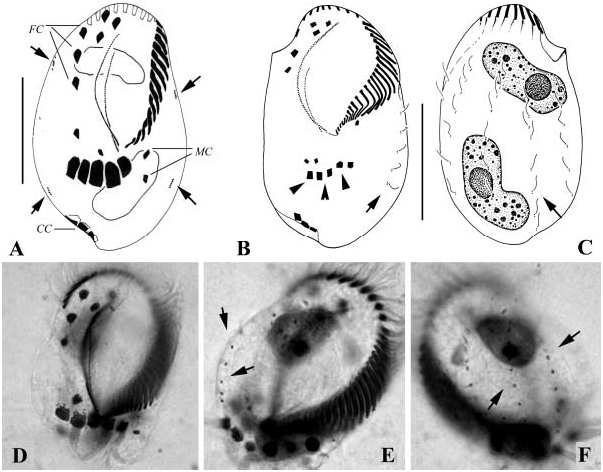
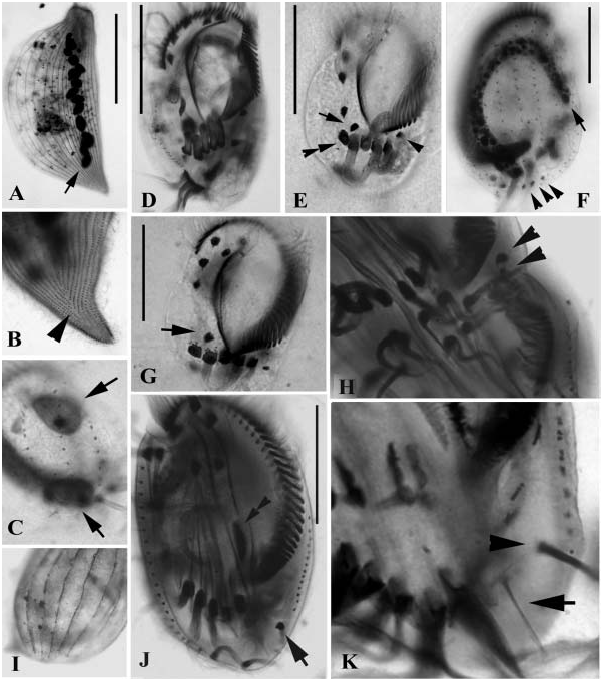
( Figures 12A–F View Figure 12 )
The present population corresponds perfectly the type population ( Wilbert and Song, 2005). Hence it is not necessary to make a further description. We provide only a few comments.
In total three species have been reported in the genus Hemigastrostyla ( Song and Wilbert 1997; Wilbert and Song, 2005). This organism differs from the closelyrelated H. enigmatica ( Dragesco and Dragesco-Kernéis, 1986) in its lower number of frontal and buccal cirri (eight versus more than eight in the latter), no ‘‘extra’’ ventral cirri (versus present), fewer dorsal kineties (three versus five) and the body shape (non-cephalized versus cephalized) ( Dragesco and Dragesco-Kernéis 1986; Song and Wilbert 1997). It differs from H. stenocephala ( Borror 1963) in conspicuously shorter dorsal cilia (ca. 3 Mm versus 16 Mm) and fewer frontal and ventral cirri (13 versus 17 in H. stenocephala ) ( Borror, 1963).
Diophrys appendiculata (Ehrenberg, 1838)
( Figures 5A View Figure 5 , 10E View Figure 10 )
The infraciliature of the current population shows typical characters, as were redescribed by Song and Packroff (1997) and Song et al. (2007) ( Figures 5A View Figure 5 , 10E View Figure 10 ). Hence the identification is quite certain.
This form had been frequently confused with D. oligothrix in the literature simply because there was no clear outline for either of then. According to the redefinition by Song and Packroff (1997), this organism can be distinguished from the closely-related D. oligothrix by the following criteria: (1) smaller size (usually about 60 Mm vs. mostly.80 Mm in body length); (2) broadly oval body shape and dominant buccal field, and (3) arrangement of cilia in dorsolateral kineties (in broken pattern vs. continuous in the latter).
Diophrys aff. oligothrix Borror, 1965
( Figures 5B–F View Figure 5 ; 10C, G View Figure 10 )
The well-known D. oligothrix was redefined by Song and Packroff (1997), Chen and Song (2002) and Song et al. (2007), was also isolated in the Antarctic area ( Song and Wilbert 2002a) and was incorrectly identified as another Antarctic form in the description by Petz et al. (1995). Considering all morphological features, the present population should be conspecific with the morphotype reported by Petz et al. (1995). Since some important in vivo features remain unavailable, we have to treat this form as an uncertain form here and wait for further investigations.
Description
It is likely that several living features in this organism are rather population- or even individual-dependent, like the size (60–120 Mm in length), body shape (slender to broadly oval), the number of dorsal kineties (4–5, mostly five) etc., an attribute shared with other related congeners, especially with D. oligothrix and D. appendiculata ( Song and Packroff 1997; Song and Wilbert 2002a).
Remarks
This organism differs from D. oligothrix in consistently having only a single marginal cirrus, a lower number of dikinetids but longer cilia in dorsal kineties, relatively shorter ciliary organelles (e.g. caudal cirri) and mostly more oval body shape ( Song and Packroff 1997). Compared with the closely-related D. appendiculata (see above), the present form, like D. oligothrix , can be identified by the combination of the following characters: (1) continuous ciliary rows on dorsal side with loosely arranged cilia; (2) usually four dorsal kineties and about 30 membranelles; (3) body generally slender and usually c. 80 Mm long in vivo; (4) often prolonged anterior membranelles and caudal cirri, and (5) buccal field usually about half of body length. In addition, this species is often slightly yellowish in colour when observed at low magnifications.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Holosticha diademata (Rees, 1884)
| Wilbert, Norbert & Song, Weibo 2008 |
Hemigastrostyla szaboi
| Wilbert and Song 2005 |