
Caligus hyporhamphi, Boxshall, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4398.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:79E3EB78-D1C3-45CF-AB13-F8E61C936252 |
|
DOI |
https://doi.org/10.5281/zenodo.5952170 |
|
persistent identifier |
https://treatment.plazi.org/id/03B587F2-AA2F-4D62-B6F8-FA923A3DFD34 |
|
treatment provided by |
Plazi |
|
scientific name |
Caligus hyporhamphi |
| status |
sp. nov. |
Caligus hyporhamphi sp. nov.
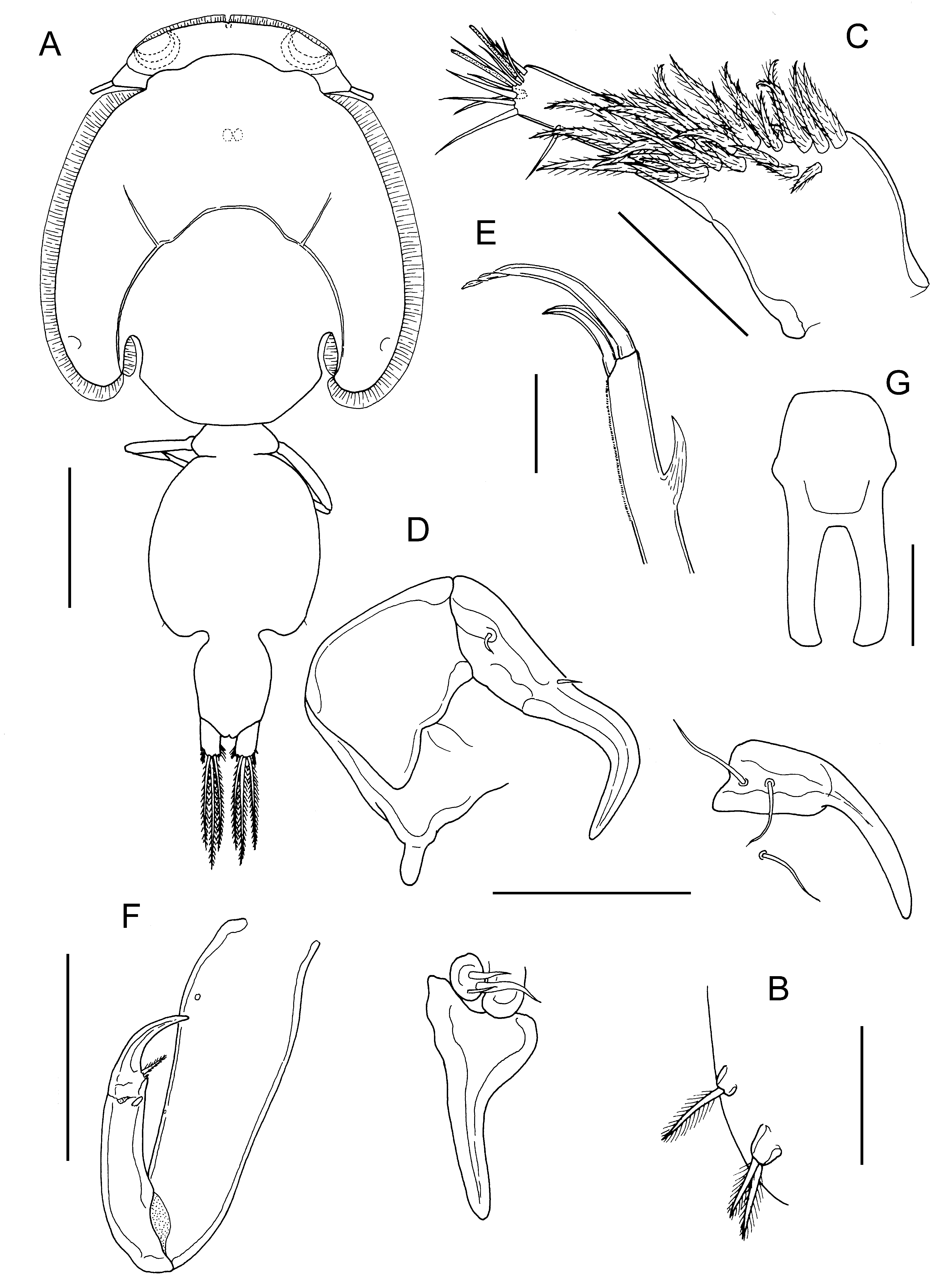
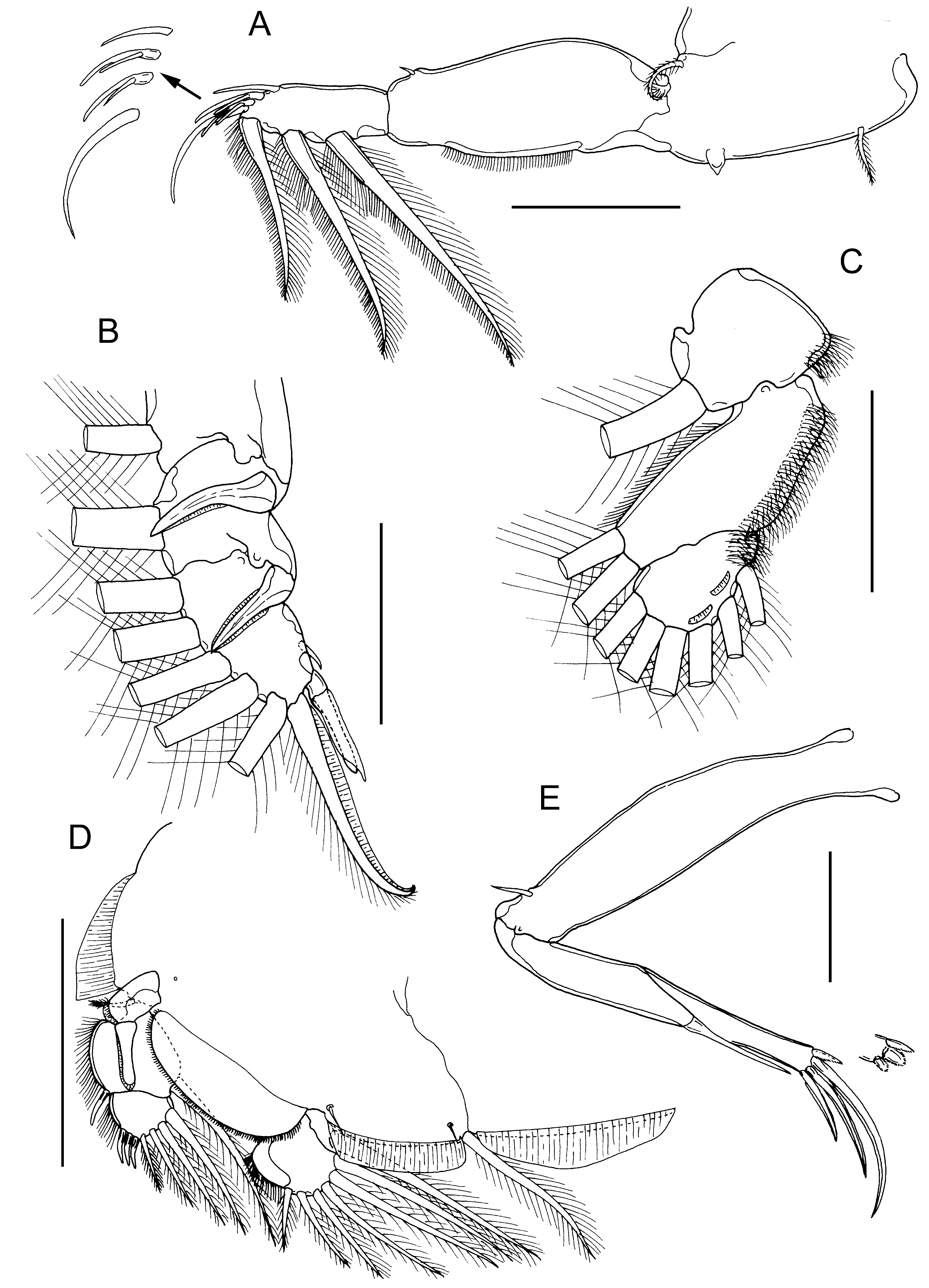
( Figs. 30 View FIGURE 30 , 31 View FIGURE 31 )
Type material. Holotype ♀ from Hyporhamphus quoyi (Valenciennes, 1847) (TC17880) 5 July 2016, QM Reg. No. W53074.
Type Host. Hyporhamphus quoyi (Valenciennes, 1847) .
Site on host. Unknown (in body wash).
Etymology. The species name refers to the generic name of its type host.
Description. Holotype female ( Fig. 30A View FIGURE 30 ) body length 5.26 mm, including caudal rami. Cephalothorax slightly longer than wide (2.90 x 2.48 mm). Free margin of thoracic portion of dorsal cephalothoracic shield extending posteriorly beyond rear margins of lateral portions. Large lunules present ventrally on frontal plates. Genital complex just longer than wide (1.29 x 1.23 mm), with convex lateral margins and rounded corners ( Fig. 30A View FIGURE 30 ); fifth legs ( Fig. 30B View FIGURE 30 ) located ventrally close to posterolateral corners. Genital complex about 1.5 times longer than abdomen. Abdomen 1-segmented, about 1.6 times longer than wide (0.87 x 0.55 mm) in ventral view; carrying paired caudal rami distally; anal slit terminal. Caudal rami with parallel sides, about 1.4 times longer than wide, measured at midpoints of margins: each ramus armed with short hirsute seta at inner distal angle, slightly longer hirsute seta at outer distal angle, minute hirsute seta located just ventral to outer distal seta, and 3 long plumose setae on distal margin.
Antennule ( Fig. 30C View FIGURE 30 ) 2-segmented; large proximal segment with 25 plumose setae arrayed along anteroventral surface plus 2 setae located dorsally; distal segment bearing 12 elements (10 setae plus 2 aesthetascs) around apex, plus isolated seta on posterior margin. Antenna ( Fig. 30D View FIGURE 30 ) comprising proximal segment bearing narrow, posteriorly-directed spinous process; middle segment subrectangular, unarmed; terminal segment forming recurved claw bearing seta on small swelling proximally and distal seta near anterior margin. Post-antennal process welldeveloped, slender and slightly curved; ornamented with 2 unisensillate papillae on basal part and single similar unisensillate papilla on adjacent ventral cephalothoracic surface ( Fig. 30D View FIGURE 30 ).
Mandible of typical stylet-like structure, with 12 marginal teeth. Maxillule ( Fig. 30D View FIGURE 30 ) comprising anterior papilla bearing 3 unequal, naked setae and slender posterior, tine-like process; sclerite adjacent to anterior papilla with rounded posterior process overlapping base of posterior process. Maxilla 2-segmented ( Fig. 30E View FIGURE 30 ), comprising elongate syncoxa and basis; basis bearing membranous but spiniform subapical flabellum on anterior margin, ornamented with minute corrugations distally along posterior margin, and terminating in 2 claw-like elements (calamus and canna): calamus longer than canna, ornamented with strips of serrated membrane arranged obliquely around surface; canna ornamented with bilateral strips of serrated membrane. Maxilliped subchelate ( Fig. 30F View FIGURE 30 ); slender protopodal segment lacking any process on myxal surface; distal subchela with apical claw separated from proximal segmental part by incomplete suture; seta present on concave margin.
Sternal furca ( Fig. 30G View FIGURE 30 ) with long parallel tines, slightly expanded and incurved at tip.
First swimming leg pair ( Fig. 31A View FIGURE 31 ) joined by slender intercoxal sclerite; sympod with inner and outer plumose setae; endopod represented by unarmed tapering process on posterior margin of basis. Exopod 2-segmented; directed laterally and forming main axis of leg; first segment robust, about 2.4 times longer than wide armed with small outer (anterior) spine, and ornamented with row of setules along posterior margin; second segment armed with 3 long plumose setae along posterior margin and 4 distal elements along oblique distal margin. Distal elements as follows: spine 1 (anterior-most) simple, about as long as spine 2; spines 2 and 3 subequal in length, each with accessory process; seta 4 about twice as long as spines and slightly longer than segment.
Second leg biramous, with flattened protopodal segments and 3-segmented rami. Coxae of leg pair joined by intercoxal sclerite bearing marginal membrane posteriorly. Coxa with plumose seta and surface sensilla. Basis armed with outer naked seta; ornamented with surface sensilla, marginal membrane posteriorly, and flap of membrane anteriorly, reflexed back over dorsal surface of segment. Exopodal segments 1 and 2 ( Fig. 31B View FIGURE 31 ) each with large outer spines reflexed and extending obliquely across ventral surface of ramus, plus inner plumose seta; first segment with flap of membrane anteriorly reflexed back across dorsal surface of ramus; segment 3 with 2 outer spines (proximal spine minute, distal spine armed with flap of membrane), apical spine with marginal membrane laterally and pinnules medially, and 5 inner plumose setae. First and second endopodal segments ( Fig. 31C View FIGURE 31 ) armed with 1 and 2 inner plumose setae, respectively; segment 3 armed with 6 plumose setae and ornamented with 2 shallow crescentic strips of membrane; outer margins of all endopodal segments ornamented with fine setules extending from margin onto surface of segment.
Third leg pair ( Fig. 31D View FIGURE 31 ) forming flattened plate closing posterior part of cephalothoracic sucker as typical for genus. Protopodal part flattened and joined by plate-like intercoxal sclerite, forming apron ornamented with marginal membrane posteriorly and along lateral margin anterior to exopod: bearing inner plumose seta at junction with intercoxal plate, and outer plumose seta dorsal to base of exopod; long sensillae located adjacent to inner coxal seta and adjacent to origin of endopod. Exopod 3-segmented; first segment armed with straight outer claw directed over ventral surface of ramus, claw and outer distal margin of segment ornamented with strip of membrane; second segment with slender outer spine and inner plumose seta; third with 3 outer spines and 4 inner plumose setae; outer margins of segments 2 and 3 ornamented with row of slender setules. Endopod 2-segmented; first segment forming flap-like velum ornamented with row of fine setules along free margin, closing space between rami, and armed with inner plumose seta; compound distal segment expanded laterally, armed with 6 setal elements increasing in length from outermost to innermost.
Fourth leg ( Fig. 31E View FIGURE 31 ) 3-segmented, comprising slender protopodal segment and 2-segmented exopod: protopodal segment armed with plumose seta distally; first exopodal segment armed with long slender outer spine; compound distal segment with 3 distal margin spines increasing in size from outer to inner; innermost with elongate pecten at base.
Fifth legs located posterolaterally on genital complex ( Fig. 30B View FIGURE 30 ); each fifth leg comprising anterior papilla bearing plumose seta (representing outer protopodal seta) and inner exopodal papilla armed with 2 plumose setae.
…… continued on the next page These poorly described species are included but many morphological details are lacking.
Remarks. The female from H. quoyi has a 3-segmented leg 4 with the first and second exopodal segments bearing 1 and 3 distal spines, respectively. This configuration, where there is no lateral spine on the compound second segment, is exhibited by about 65 valid species of Caligus . In 1980, Boxshall & Gurney recognized a cluster of 28 species, which they referred to as the C. macarovi -group, which was characterized by this form of leg 4 exhibited in combination with a suite of other features including: the distal exopodal segment of leg 1 armed with 3 posterior margin plumose setae, spines 1, 2 and 3 all subequal in length and only spines 2 and 3 with accessory processes, and seta 4 markedly longer than the spines, plus a 1-segmented abdomen in the female. In addition, they noted that many core members of this group also possessed serrations distally on the margin of the brachium of the maxilla. Numerous additional species sharing most or all of these character states have been described since 1980, therefore all 42 current members of the C. macarovi group are listed in Table 5. Excluded are another 23 species which have a 3-segmented leg 4 with the first and second exopodal segments bearing 1 and 3 distal spines, but which display a different form of armature on the distal exopodal segment of leg 1, including: having spine 1 reduced (e.g. C. cresseyorum ), seta 4 shorter than the spines (e.g. C. pampi ), lacking plumose setae on posterior margin (e.g. C. lethrinicola ), possessing an accessory process on spine 1 (e.g. C. amblygenitalis ), or lacking an accessory process on spine 2 and/or spine 3 (e.g. C. calotomi ).
The genital complex in female caligids can vary in shape somewhat according to the reproductive state of the individual female ( Parker et al., 1968; Boxshall, 1974) and it is necessary to allow for potential variability when making comparisons of body proportions between species. The new species has a genital complex that is about as long as wide: allowing for about variability of about 10% eliminates species with a genital complex that is markedly wider than long (length: width ratio ź 0.8:1) and species where it is markedly longer than wide (L: W ratio ± 1.2:1). Species exhibiting these shapes are delimited in Table 5, which lists species in order from a “broad” to an “elongate” genital complex.
Only 18 species share a genital complex with a L: W ratio between 0.9:1 and 1.1:1. Four of these, C. absens , C. hamruri , C. thyrsitae and C. longiabdominis , have an abdomen that is at least twice as long as wide, compared to 1.4 times in the new species. Another seven species, C. aduncus , C. antennatus , C. pseudokalumai , C. rogercresseyi , C. scribae , C. stokesi and C. tetrodontis , have a genital complex that is at least 2.4 times longer than the abdomen, compared to 1.7 times in the new species.
The remaining seven species can be compared in more detail. Caligus dampieri is an Australian species found on single host species, Acanthopagrus latus , caught at Port Samson in Western Australia (Byrnes, 1987). Both of these species share the presence of a pair of crescent-shaped membranes on the surface of the distal endopodal segment of leg 2.The most obvious difference between these two species is the wide gape between the divergent tines of the sternal furca in C. dampieri compared to the parallel to slightly convergent tines in the new species. The two segments of the exopod of leg 4 are subequal in the new species whereas in C. dampieri the second segment is markedly longer than the first. An additional minor difference is the size of the posterior process on the antenna.
Caligus itacurussensis was found on Atherinella brasiliensis (Quoy & Gaimard, 1825) (as Xenomelaniris brasiliensis ) in Sepetiba Bay, Brazil ( Luque & Cezar, 2000). It differs from the new species in having relatively shorter tines on the post-antennal process and the maxillule. The tines on the sternal furca are also shorter and are distinctly divergent rather than parallel, as in the new species. In addition the first exopodal segment of C. itacurussensis has a slightly curved outer spine and lacks marginal membrane, whereas the new species has a straight spine and the outer margin is ornamented with membrane.
Caligus latus was also found at Port Samson on Acanthopagrus latus (Byrnes, 1987) . Comparison is handicapped by lack of information on some of the appendages but the female genital complex is 4.0 times wider than the abdomen in C. latus but only 2.2 times wider in the new species. The gape between the divergent tines of the sternal furca in C. latus is very wide compared to the parallel to slightly convergent tines in the new species.
Shiino’s (1959) redescription of C. macarovi shows an elongate abdomen that is 1.9 times longer than wide, compared to only 1.4 times in the new species. The exopodal segments of leg 4 are stouter in C. macarovi and the outer spines are shorter relative to the apical spine in C. macarovi than in the new species. The posterior process on the proximal segment of the antenna and the tines on both the post-antennal process and the maxillule are all shorter and stouter in C. macarovi than in the new species.
Caligus wilsoni is a replacement name proposed by Delamare Deboutteville & Nuñes-Ruivo (1958) to accommodate material misidentified as C. belones Krøyer, 1863 by C.B. Wilson (1905). Comparison with the redescription by Cressey (1991) reveals significant differences in the shape of the sternal furca and in the armature of leg 2. The short tines of the sternal furca are broad at the base and taper towards the tip whereas in the new species the tines are elongate and do not taper distally. The outer spines on the first and second exopodal segments of leg 2 are directed nearly parallel with the long axis of the ramus in C. wilsoni but are reflexed obliquely across the surface of the ramus in the new species.
The recently established C. solea is known only from Solea solea (Linnaeus, 1758) in the Mediterranean Sea (Demirkale et al., 2014). In C. solea the distal segment of the exopod of leg 4 is markedly longer than the proximal whereas the two segments are subequal in the new species. In addition, the spines on this leg in C. solea are much shorter and more blade-like than in the new species.
Shiino (1954c) established C. triangularis to accommodate material collected from Halichoeres poecilopterus (Temminck & Schlegel, 1845) in Japanese waters. It differs from the new species in having relatively short outer spines on the exopodal segments of leg 4: the spine on the proximal exopodal segment reaches only halfway to the tip of the ramus whereas in the new species it almost reaches the tip, and the middle spine on the distal segment is less than half the length of the apical spine compared to 75% in the new species. Spines 1 to 3 are stout in C. triangularis but slender in the new species. These detailed comparisons justify the establishment of a new species to accommodate the female from Hyporhamphus quoyi .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |