
Proceratophrys redacta, Junior, Mauro Teixeira, Amaro, Renata Cecília, Recoder, Renato Sousa, Vechio, Francisco Dal & Rodrigues, Miguel Trefaut, 2012
|
publication ID |
https://doi.org/10.5281/zenodo.214281 |
|
DOI |
https://doi.org/10.5281/zenodo.6167503 |
|
persistent identifier |
https://treatment.plazi.org/id/03B68247-DD48-FD15-F78E-F9538EEFDD01 |
|
treatment provided by |
Plazi |
|
scientific name |
Proceratophrys redacta |
| status |
sp. nov. |
Proceratophrys redacta sp. nov.
( Figs. 1–3 View FIGURE 1 View FIGURE 3 )
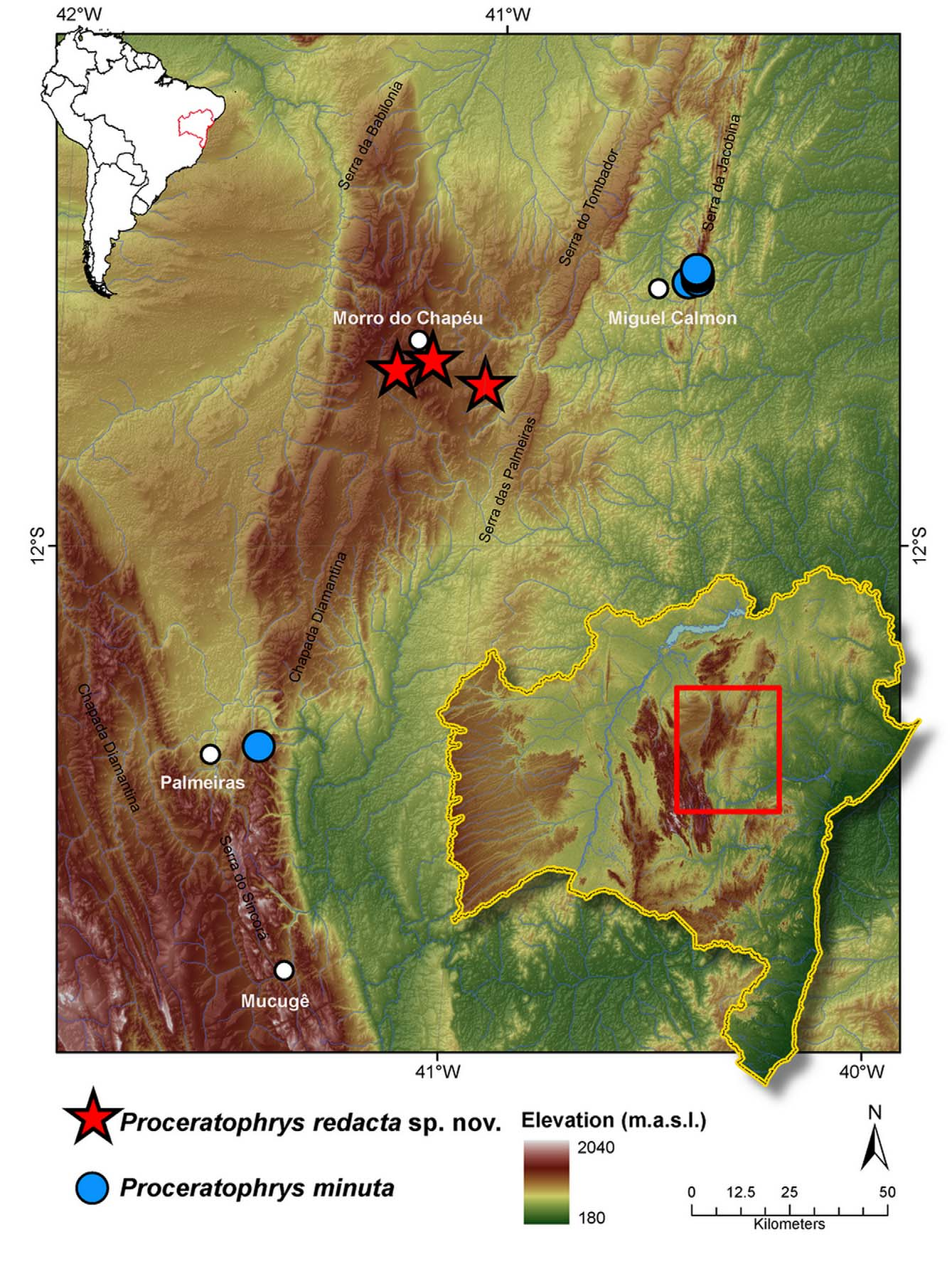
Holotype. MZUSP 150266, an adult male, field number MTR 22579, collected at Morro do Chapéu hill, Morro do Chapéu municipality, Bahia, Brazil ( 11°35'26.02"S, 41°12'32.87"W, 1254 m a.s.l. datum WGS-84), by M. Teixeira Jr, F. Dal Vechio, I. Prates, R. P. Damasceno and M. T. Rodrigues, on 30th December 2011.
Paratopotypes. MZUSP 150263, 150264, 150265, 150268, 150269, adult males, field numbers MTR 22474, 22554, 22564, 22582, 22583; MZUSP 150267, adult female, field number MTR 22580. Collected between 27th and 30th December 2011.
Paratypes. MZUSP 150272, an adult male, field number MTR 22628; MZUSP 150270, 150271, 150273 adult females, field numbers MTR 22623, MTR 22626, MTR 22629. All from an old road ( 11°33'58.3"S, 41°07'32.3"W, 929 m a.s.l., datum SAD69), at Morro do Chapéu municipality, close to city town. Collected between 31st December 2011 and 2nd January 2012.
Diagnosis. A Proceratophrys species (nuptial pads absent, enlarged glands absent, fingers webbing absent, supernumerary tubercles on hands and feet present, dorsal surfaces of fingers and toes wrinkled, and dorsum with sinuous longitudinal ridges outlining a spear-shaped ornamentation) characterized by: (1) small size ( 26.9–29.7 mm of SVL in males; 33.6–34.9 mm of SVL in females); (2) no rostral appendage; (3) no palpebral appendage; (4) snout rounded from above and vertical in profile; (5) rostral ridge sharp, slightly curved; (6) presence of interrupted sagittal ridges of warts from eyelids to sacral region; (7) absence of postocular swellings; (8) tympanic region indistinct; (9) dorsal coloration lacking distinct dark ocelli.
Comparisons with other species (data for the species in comparison are given in parenthesis). Proceratophrys redacta sp. nov. can be promptly distinguished from all species of Proceratophrys , with the exception of P. avelinoi , P. minuta , P. m o r a t o i and P. palustris , by its smaller body size, SVL ranging from 26.9–29.7 mm in males, and 33.6–34.9 mm in females ( P. appendiculata : 40.4–59.9 males, 33.5–63.2 females; P. aridus : 32.5–42.8 males, 43.2–46.3 females; P. bigibbosa : 35.5–43.8 males, 51.2–53.4 females; P. b o i e i: 39.8–61.9 males, 40–74.3 females; P. brauni : 30–34.6 males, 38.9–39.8 females; P. caramaschii : 42.7–56.3 males, 53.8– 65 females; P. concavitympanum : 39.6–57.1 males, 53.6–60.3 females; P. cristiceps : 39.5–50.2 males, 43–53.8 females; P. cururu : 36.5–43.1 males, 47–53.9 females; P. g o y a n a: 39.6–53.5 males, 54.6–60.0 females; P. laticeps : 59.5– 78 males, 50.3–82.7 females; P. melanopogon : 34.9–49.9 males, 40.7–62.1 females; P. moehringi : 59.2–62.6 males, 63.6 the only female known; P. p a v i o t t i: 41.9–53.2 males, 50.9–51.8 females; P. phyllostomus : 55.4 the only male known, 40.9–76.6 females; P. re n a l i s: 36.8– 53 males, 38.5–71.9 females; P. rondonae : 62.9 the only male knwon; P. sanctaritae : 38.4–45.5 males, 60.8 female; P. schirchi : 31.6–40.4 males; P. strussmanae : 41.1–47.3 males, 52.6–59.8 females; P. subguttata : 36.7–41.8 males, 36.6–58.9 females; P. tupinamba . 52.6–63.4 males, 37.7–72.8 females; P. vielliardi : 39.1–41.9 males, 45.5 the only female known; P. re d a c t a sp. nov. can be distinguished from P. minuta by its larger size, SVL ranging from 26.9–29.7 mm in males, and 33.6–34.9 mm in females, by having an incomplete sagittal ridge of warts extending from eyelid to sacral region, and having a row of small, rounded tubercles in the eyelid (SVL 19.2–25.2 mm males, 25.9–31.9 mm females; presence of a complete sagittal ridge of warts from eyelids to urostile; a row of enlarged, pointed tubercles in the eyelid is present); from P. avelinoi , P. palustris , P. brauni and P. bigibbosa , it can be distinguished by lacking postocular swellings (present) a additionally from P. avelinoi and also P. schirchi by having a row of small, rounded tubercles in the eyelid (a row of enlarged, pointed tubercles in the eyelid is present); from P. moratoi , P. palustris and P. vielliardi it can be distinguished by having the dorsal and lateral surfaces of body covered with small granules with a few scattered larger tubercles, incomplete sagittal ridge of tubercles extending from eyelid to sacral region, and dorsal coloration variable, but never with dark irregular ocelli (dorsal surfaces with many large tubercles, no sagittal ridge of tubercles and dorsal coloration with irregular ocelli).
Additionally P. re d a c t a sp. nov. can also be distinguished from P. appendiculata , P. boiei , P. laticeps , P. melanopogon , P. moehringi , P. p a v i o t t i, P. phyllostomus , P. re n a l i s, P. rondonae , P. sanctaritae , P. subguttata , and P. tupinamba by lacking a single long horn-like palpebral appendage (present); from P. appendiculata , P. melanopogon , P. laticeps , P. phyllostomus , P. subguttata and P. tupinamba by lacking a rostral appendage (present); and from P. concavitympanum and P. strussmannae by having an indistinct tympanic region (tympanic region defined by a concave depression on the skin).
2. Details of the head in dorsal view (A), and profile (B), and ventral views of right hand (C) and foot (D) of the holotype (MZUSP 150266) of Proceratophrys redacta sp. nov. Scale bars correspond to 5 mm.
Description of holotype. Body small, stout, nearly ovoid ( Fig. 1 View FIGURE 1 ); 28.3 mm in SVL. Head wider than long; head length 87% of head width. Snout rounded in dorsal view, vertical to slightly oblique in profile (Fig. 2). Upper eyelids without appendages, covered with a row of small rounded tubercles. Canthal crests distinct, loreal region slightly concave. Interocular crests absent. Interorbital distance 88% of upper eyelid width, 23.9% of head width. No oblique rows of warts on head sides. Three prominent and yellowish postrictal tubercles posteriorly to mouth commissure. Pre-ocular crests present, short and vertical. Eyes large, eye diameter 34% of head length, and 1.52 times the eye to nostril distance. Tympanum hidden. Nares prominent, nostrils elliptical, nearly horizontal in profile, diverging posteriorly in dorsal view. Tongue cordiform, free posteriorly. Vomerine teeth in two short rows between choannae. Choannae large, rounded, separated by a distance of about 2.5 times its diameter. Vocal sac single, subgular; vocal slits present, lateral to tongue on the floor of mouth. Arms short and robust; finger length IV<II<I<III. Finger tips rounded, not expanded. Fingers not webbed, bordered laterally by distinct ridges of tubercles. Nuptial pads absent in males. Inner carpal tubercle elliptical. Outer carpal tubercle bifid, ovoid with a ventral invagination inwards. Subarticular tubercles large, rectangular with rounded corners; supernumerary tubercles well defined, rounded. Outer margin of forearms with enlarged tubercles, but not forming well defined rows. Legs short and moderately robust, leg length (THL+TL) 75% of SVL, foot plus tarsus 91% of SVL. Toe lengths I<II<V<III<IV. Toe tips rounded, smooth, without expansions, bordered laterally by distinct ridges. Toes basally webbed, web formula I 0–1- II 1 ½–2- III 1 -–3+ IV 3 -– 1- V. Inner metatarsal tubercle large, globular, and semicircular. Outer metatarsal tubercle small, rounded. Subarticular tubercles large, rectangular with rounded corners; supernumerary tubercles present. Tarsal tubercles in a ventrolateral row, forming a lateral ridge on foot. Dorsal surfaces granulate with larger granules irregularly distributed in the dorsum, especially within dorsal ridges. Symmetrical sinuous longitudinal ridges of tubercles on dorsum, from posterior border of eyelids and scapulae, converging at middle of the body, and fading posteriorly, not reaching the urostile. Ventral surfaces shagreened. A pair of enlarged light tubercles below the anal region.
Color in preservative. Dorsal background color light brown. Dark brown interorbital bar projecting posteriorly. Dark brown triangular areas irregularly delimited along the outer margins of the dorsal sagittal crests projecting posteriorly over scapulae, curving inwards, reaching sacrum region. A small chevron medially located at the dorsum. Irregular dark brown markings distributed over the dorsum and along flanks. Limbs with transverse dark brown bars. Ventrally, ground color yellowish-brown; darker brown on palms, soles, anal region and throat. Three oblique dark brown stripes on sides of head, accompanying ridges of tubercles, and two less defined on snout to lips. Upper lip cream.
Measurements of holotype (in mm). SVL 28.3; HL 11.0; HW 12.6; ED 3.8; END 2.5; UEW 3.4; IOD 3.0; IND 2.1; THL 11.4; TL 9.9; FTL 15.9; HAL 12.4.
Variation. We found great variation on color pattern among specimens, the more common individuals have a similar pattern to that described for the holotype ( Fig. 3 View FIGURE 3 A and C) as well as a more melanic-marbled pattern ( Fig. 3 View FIGURE 3 B). Although less common, a pattern with a yellowish brown inner to the borders of the sagittal crests is also found ( Fig. 3 View FIGURE 3 D). Although males and females share the same dorsal color variation, the throat in males present a dark brown color, and females a whitish coloration. Measurements for P. re d a c t a sp. nov. and P. m i n u t a are presented in Table 1.
Etymology. The name of the new species is derived from the Latin adjective redactus that means reduced, in allusion to the small size of the new species.
Natural history and distribution. Proceratophrys redacta sp. nov. is known only from the surroundings of type locality, at the montane region of Morro do Chapéu municipality, close to where P. m i n u t a occurs (about 100 km away) ( Fig. 4 View FIGURE 4 ). The region is dominated by the Mesoproterozoic sandstones of Morro do Chapéu formation ( Battilani et al. 1996), reaching up to 1300 m a.s.l., the climate is a sub-warm semi-humid with annual mean temperatures under 22o C, (4o C was already recorded), and precipitation is regular with a short dry season ( Nimer 1972). Landscape in the area consists of an extensive and relatively flat plateau situated around 1200 m from which the hill of Morro do Chapéu (“hat hill”) emerges. At the plain sandy depositional areas grows an open shrubby vegetation, however at the richer soil areas a semi-deciduous forest is found; the bare rock at the mountain tops and eroded areas are covered by a typical rocky meadows vegetation, where Cactaceae , Bromeliaceae , and Velloziaceae are abundant. Steeper areas are covered by a lower open forest, the ground is sandy and covered with a deep layer of leaves and a few scattered small ponds are present, surrounded with mosses.
Proceratoprys redacta sp.n. Proceratophrys minuta
Males (N=6) Females (N=4) Males (N=10) Females (N=11) SVL 28.4 (26.9–29.7) 34.4 (33.5–34.9) 20.9 (19.2–22.6) 28.9 (25.9–31.6) HL 11.6 (10.8–12.4) 13.3 (12.7–14.1) 9.1 (8.6–9.5) 11.2 (10.5–12.2) HW 13 (12.2–13.8) 15.7 (15.2–16.2) 10.0 (9.2–10.8) 13.2 (12.2–14.1) ED 4.1 (3.7–4.4) 4.6 (4.5–4.7) 2.6 (2.4–2.9) 3.2 (2.9–3.6) END 2.5 (2.4–2.6) 2.9 (2.7–3.1) 2.1 (1.8–2.3) 2.6 (2.4–3.2) UEW 3.4 (3.3–3.7) 4 (3.8–4.2) 3.1 (2.8–3.4) 3.8 (3.4–4.3) IOD 3 (2.4–3.4) 3.4 (3.1–3.8) 2.5 (2.3–3.0) 3.2 (3.1–3.4) IND 1.9 (1.7–2.2) 2.1 (2–2.5) 1.8 (1.6–1.9) 2.2 (2.1–2.3) THL 12.1 (11.3–12.8) 13.9 (13.1–14.5) 9.5 (8.6–10.4) 11.9 (11.3–12.4) TL 10.2 (9.8–10.5) 12 (11.9–12.2) 8.5 (8.1–9.1) 10.9 (10.5–11.3) FTL 17.2 (15.9–18.4) 20.1 (19.7–20.4) 13.1 (11.9–14.1) 17.2 (15.8–17.8) HAL 13.3 (12.3–14.2) 23.9 (15.9–46.4) 11.1 (10.4–12.2) 14.5 (14.1–14.8) P. redacta sp. nov. was found active on the ground at night, within the montane forest and the adjacent open vegetation ( Fig. 5 View FIGURE 5 ). It was also captured in pitfall traps, along the Morro do Chapéu hill, about 1250 m a.s.l. and in the dense vegetation at the shores of streams running over bare rock outcrops, as those of Ferro Doido waterfall at about 890 m a.s.l. No individual was heard calling. Also none was found at the dry sandy scrublands, the most abundant vegetation at this area, indicating a restriction to this open mesic areas.
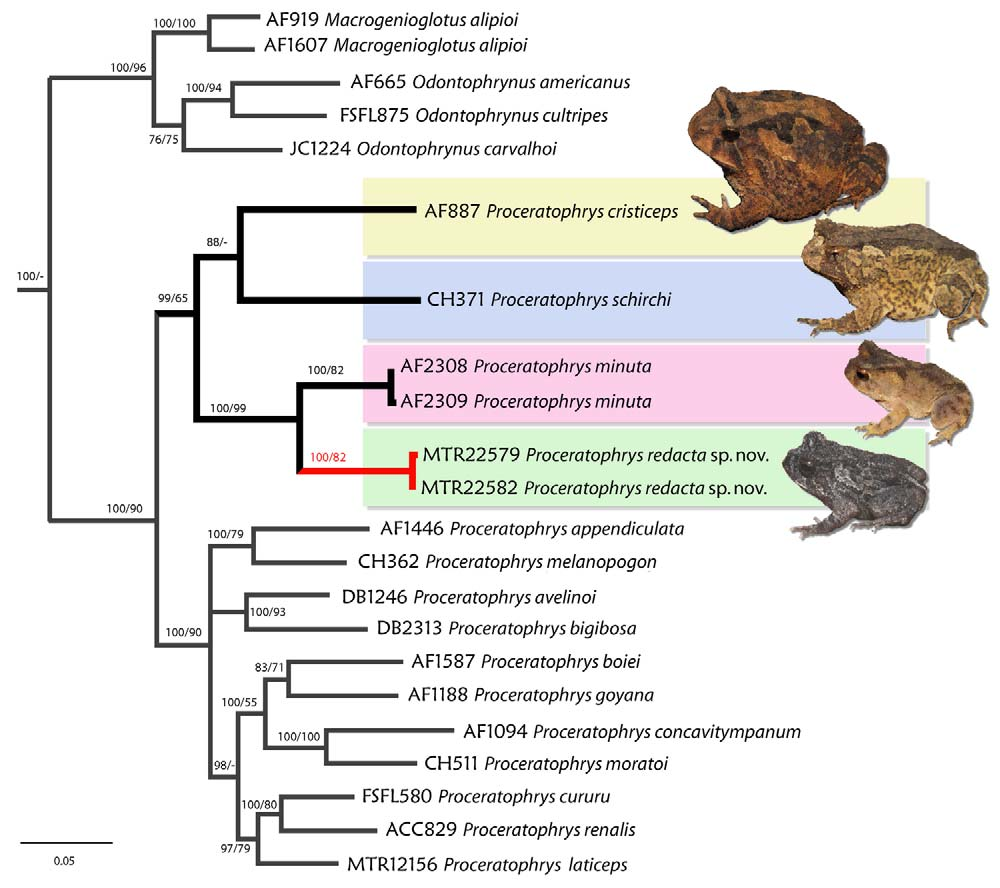
Molecular phylogeny. Our analyses recovered Proceratophrys as monophyletic, and consisting of two main clades. The first is composed by P. cristiceps , P schirchi , P. m i n u t a and P. redacta sp. nov. All analyses recovered Proceratophrys redacta sp. nov. as sister to P. m i n u t a, both being sister of a clade assembling P. cristiceps and P. schirchi ( Fig. 6 View FIGURE 6 ). The second clade is composed by the remaining species of Proceratophrys , however the intrageneric relationships are poorly resolved. Three different lineages were recovered in this clade: 1) P. appendiculata and P. melanopogon as sister species; 2) P. avelinoi and P. bigibbosa as sister species; and 3) a clade containing P. boiei , P. goyana , P. concavitympanum , P. moratoi , P. cururu , P. re n al is, and P. laticeps . In the latter Proceratophrys moratoi is sister to P. concavitympanum and both sister to P. boiei and P. goyana . This latter clade is sister to a clade where Proceratophrys laticeps is basal to P. cururu and P. re n a l i s, which are recovered as sister species.
Uncorrected p -distances ranged from 12 to 21% for cyt b, 3 to 11% for 16S and 1 to 5% for Rag-1 among Proceratophrys species (Table 2). Genetic distances between P. minuta and P. re d a c t a sp. nov. were 12-13% for cyt b, 4% for 16S and 2% for Rag-1 and are compatible with interspecific distances found among Proceratophrys species (Table 2). It is important to note that none of the differences found on Rag-1 between P. m i n u t a and P. redacta sp nov. were related to heterozygous sites.



| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |