
Ouratea pastazana Sastre & Offroy, 2009
|
publication ID |
https://doi.org/10.5252/a2009n1a5 |
|
persistent identifier |
https://treatment.plazi.org/id/03B687D1-2B5E-5310-FF1B-F98C684A9EBD |
|
treatment provided by |
Carolina |
|
scientific name |
Ouratea pastazana Sastre & Offroy |
| status |
sp. nov. |
Ouratea pastazana Sastre & Offroy View in CoL , sp. nov.
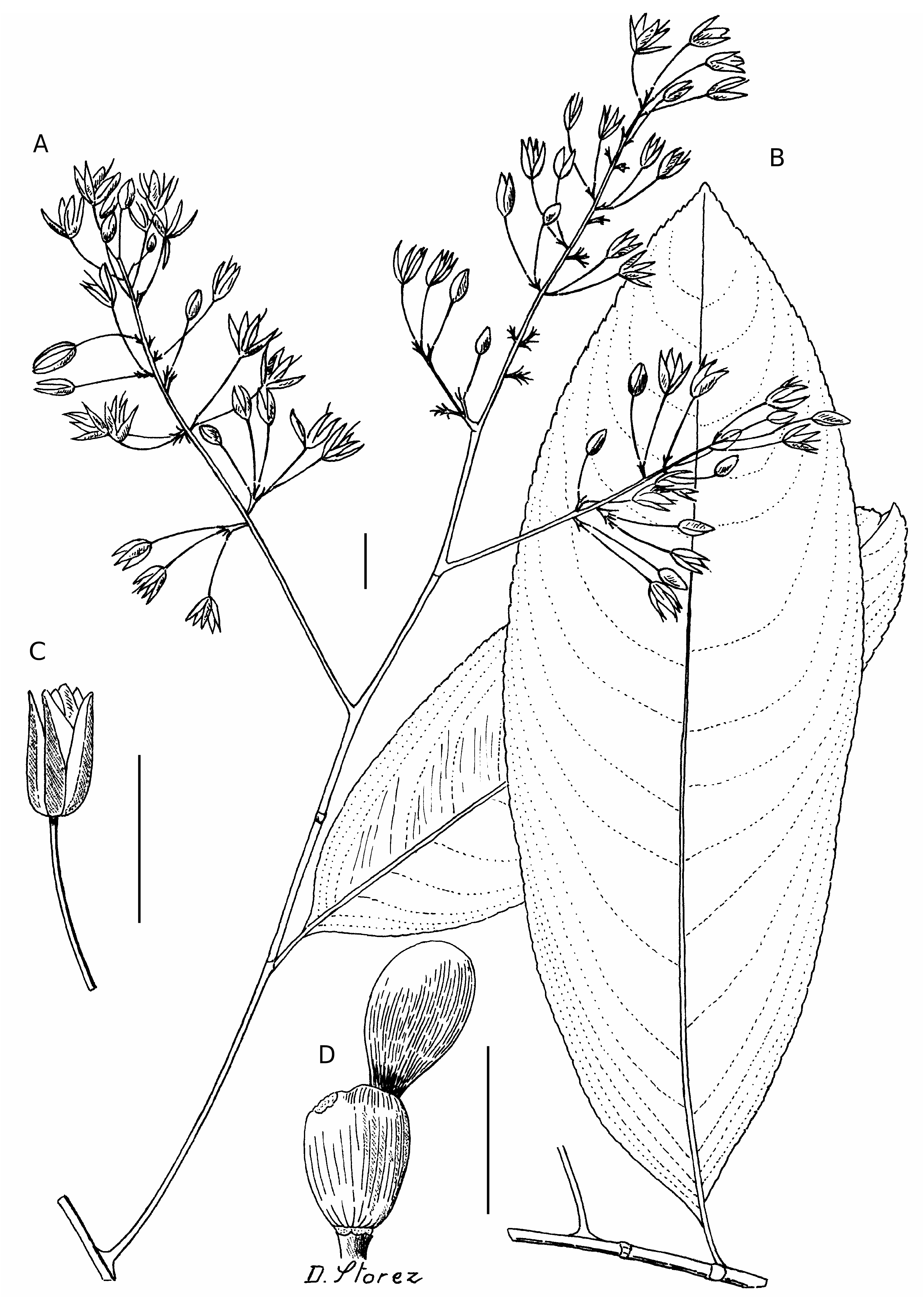
( Fig. 4 View FIG )
Ab Ouratea paraguayensi foliorum laminis membranaceis vel chartaceis, margine serrulata (vs serrata). Ab O. superba floribus, sepalis, 7,5-8 × c. 2 mm (vs (10-)12-15 × 3-4 mm), petalis 9 × 3 mm (vs 12-16 × (6-) 9-10 mm), staminis 7,5-8 mm longis (vs 10-11 mm), stylo 6-7 mm longo (vs 9-10 mm), differt. Ab O. marahuacensi foliorum laminis membranaceis vel coriaceis margine dentibus non vicinis; inflorescentiis laxis, floribus, sepalis, 7,5-8 mm longis (vs 6-7,5 mm), petalis 9 mm longis (vs 6-7,5 mm), differt. Ab O. apurensi, foliorum laminis ovatis vel ellipticis margine serratula (vs crenulata), nerviis lateralibus tantum principalibus prominulis (vs omnibus), differt.
TYPUS. — Équateur. Prov. Pastaza, Ceilàn, Pica from Ceilàn to Río Conocaco on the N side of the Río Curaray , 75°40’W, 1°36’S, alt. 200 m, flooded forest, 6. GoogleMaps VI.1980 GoogleMaps , J. Brandbyge & E. Asanza C. 31644 (holo-, AAU!).
PARATYPES. — Équateur. Prov. Pastaza, Lorocachi , a 2 horas en deslizador agues arriba, 75°59’W, 1°36’S, alt. 200 m, margin of the river, 31. V.1980, J GoogleMaps . Jamarillo , F . Coello & A . Freire 31588 ( AAU!). — Ibid., about 5 km upriver Río Curaray , 30. V.1980, J . Brandbyge & E . Asanza C . 31262 (AAU!).
DESCRIPTION
Arbre à arbuste, 3-5(-35) m de hauteur, à rameaux glabres. Feuilles à pétiole long de 3-7 mm; limbe membraneux, ovale, 10-20 × 2,5-6,5 cm, apex obtus à aigu, acuminé, marge denticulée, base atténuée, nervure principale peu saillante, nervures secondaires inégales, les plus grosses, camptodromes, arquées, rejoignant tangentiellement la marge, les plus fines, peu visibles, peu arquées, subparallèles, ne rejoignant jamais la nervure marginale, nervures tertiaires très peu visibles, droites, subparallèles et perpendiculaires avec la nervure médiane. Inflorescences axillaires ou situées au sommet de courts rameaux, panicules de 4-20 cm de longueur. Fleurs à pédoncule de 5-8 mm jusqu’à 13 mm à l’anthèse; sépales, 5, libres persistant après l’anthèse, coriaces, ovés, 7-8 × c. 2 mm, sommet aigu à mucroné; pétales, 5, libres, membraneux, c. 10 × 4 mm, sommet obtus; étamines, 10, libres, 9-10 × c. 1 mm; carpelles, 5, soudés à la base, 1 mm de hauteur, style, 5-7 mm de longueur. Fruits du type « glandularium », sépales caduques parfois persistants, torus conique, 6-8 × 5 mm, parfois arqué, carpide 1, fusiforme à sphérique, 5-8 × 5 mm.
REMARQUES
Ouratea pastazana diffère d’ O. paraguayensis par ses feuilles membraneuses à marges serrulées et
non serretées, d’ O. superba essentiellement par ses fleurs plus petites avec ses sépales de 7,5-8 × c. 2 mm ( vs (10-)12-15 × 3-4 mm), ses pétales de 9 × 3 mm ( vs 12-16 × (6-) 9-10 mm), ses étamines de 7,5-8 mm de longueur ( vs 10-11 mm) et son style de 6-7 mm de longueur ( vs 9-10 mm). Il diffère aussi d’ O. marahuacensis par ses feuilles membraneuses à marges foliaires à petites dents non serrées, par ses fleurs légèrement plus grandes avec des sépales de 7,5-8 mm de longueur ( vs 6-7,5 mm) et ses pétales de 9 mm de longueur ( vs 6-7,5 mm). Il diffère d’ O. apurensis , par ses feuilles ovales et non elliptiques à marges serrulées et non crénelées et ses nervures latérales où seules les principales sont saillantes, alors qu’elles sont toutes nettement saillantes chez O. apurensis ( Tableau 1).
CONSIDÉRATIONS BIOGÉOGRAPHIQUES
Contrairement à de nombreuses espèces d’ Ouratea , O. superba décrite du Guyana ( Engler 1876), possède une aire assez vaste incluant d’une part, le Suriname, la Guyane vénézuélienne et le cours supérieur de l’Orénoque, et d’autre part, le cours de l’Amazone (sous tous ses noms) avec des affluents majeurs, le Río Negro jusqu’au canal Casiquiare faisant la jonction avec l’Orénoque, le Río Putumayo de la Colombie jusqu’en Équateur et le Río Caquetá dans sa partie colombienne. Sur une aire aussi vaste, elle a été baptisée et décrite de nombreuses fois d’où la relative importance de sa synonymie. Il semble logique que cette espèce fondamentalement ripicole, y compris en bordure de lagunes puisse avoir une aire aussi importante, les graines étant facilement transportées par les eaux.
Par contre, les quatre espèces affines à petites fleurs, également ripicoles, possèdent des aires nettement plus restreintes qui se situent en périphérie de celle d’ O. superba . (Fig. 5). Ainsi O. marahuacensis n’a été récoltée qu’au Venezuela dans l’état d’Amazonas, au Cerro Marahuaca, à 1150 m d’altitude ( Maguire & Steyermark 1989). Ouratea apurensis , aussi du Venezuela ( état d’Apure), dans les « llanos » vénézuéliens situés près de la frontière avec la Colombie, n’est connue que de la «Reserva Forestal San Camilo», à 250-280 m d’altitude ( Sastre 1995). Ouratea pastazana n’est connue que de l’Équateur, de la province de Pastaza, aussi à basse altitude ( 200 m). Enfin O. paraguayensis possède une aire située d’une part, au Paraguay, le long du cours du Río Paraguay et de quelques affluents venant des montagnes qui marquent la frontière avec le Brésil, et d’autre part en Bolivie, le long d’affluents qui vont vers l’Amazone.
Ces quatre espèces se distinguent nettement d’ O. superba par la taille des fleurs. Par contre, les caractères pour bien les différencier entre elles sont plus subtils ( Tableau 1). Ces espèces sont globalement situées dans une aire péri-amazonienne proche de celle du type I décrit par de Granville (1992). Ces remarques pourraient suggérer une origine commune suivie d’une fragmentation d’aires.
Aujourd’hui, après les recherches de Turner (1965) sur les papillons amazoniens, de Haffer (1967) sur les oiseaux, de Vanzolini & Williams (1970) sur les reptiles et de Prance (1973) sur diverses familles botaniques, on admet la possibilité de changements climatiques en Amérique intertropicale, avec alternances de périodes sèches et de périodes humides. Ceci était en contradiction avec les concepts de Darlington (1957) qui pensait qu’un climat stable avait régné durant le Pléistocène sur les tropiques américains, contrairement à ce qui avait été suggéré pour l’Afrique depuis Christ (1892). Ce dernier a été relayé dans cette pensée par divers auteurs dont Chevalier (1928), Aubréville (1938, 1962), Schnell (1976, 1987). En Amérique tropicale, les observations ont surtout concerné les milieux forestiers. En 1976, Sastre a émis l’hypothèse que des lieux non forestiers comme les inselbergs de Guyane, seraient des refuges de végétation xérophile, suivant en cela les observations d’Aubréville (1962) en Afrique. Cette idée a été élargie à la faune par Brown & Benson (1977) et Lourenço (1987).
Durant le Pléistocène, il s’est donc établi des îles écologiques (et même des archipels, Sastre 2001a) qui ont varié selon les changements climatiques: îlots forestiers en périodes sèches, îlots savanicoles en périodes humides ( Sastre 1994a).
Les fleuves (par l’apport des affluents) et les sommets montagneux (grâce à la nébulosité) auraient
110°W 100° 90° 80° 70° 60° 50° 40°
30°N
20°
10°
0°
10°
20°S
FIG. 5. — Distribution géographique d’ Ouratea paraguayensis f. paraguayensis (▲), d’ O. paraguayensis f. boliviensis (☆), des spécimens intermédiaires entre les deux formes (★), d’ O. pastazana (●), d’ O. apurensis (□), d’ O. marahuacensis (■) et d’ O. superba (zone entourée).
toujours gardé une certaine humidité et auraient donc été des refuges de flores forestières et ripicoles. Ceci pourrait expliquer le maintien d’ O. superba sur une aire aussi vaste. Par contre, les espèces à petites fleurs, en zone péri-amazonienne ont dû se trouver isolées. D’après Hernandez-Camacho (1992) et Tricart (1993), il semblerait que les «llanos » colombiens et vénézuéliens aient connu des périodes de désertification durant le Quaternaire, y compris il y a 18 000 ans lors du dernier âge glaciaire. Des pôles humides ont pu se maintenir dans les hauteurs du Venezuela dans l’état d’Amazonas comme au Cerro Marahuaca, et dans les marécages situés le long de certains affluents de l’Orénoque, expliquant ainsi la différenciation d’ O. marahuacensis et d’ O. apurensis , devenues espèces endémovicariantes au sens de Gaussen & Leredde (1949), ceci s’ajoutant à d’autres cas de vicariance chez les Ochnacées guyanaises ( Sastre 1992, 2001b). L’Orénoque qui matérialise la frontière entre la Colombie et le Venezuela, partage deux territoires dont la géomorphologie est différente de part et d’autre. Du côté vénézuélien, dans l’arrière-pays montagneux, des sommets culminent à plus de 2600 m avec le Cerro Duida. Par contre, du côté colombien, s’étend la vaste plaine des « llanos ». De ce fait, l’Orénoque constitue une vraie frontière phytogéographique ( Sastre 1994b).
Ouratea pastazana situé dans le piémont andin équatorien dans un refuge forestier reconnu par Prance (1982) a pu profiter de conditions climatiques similaires le long de rivières s’écoulant vers l’Amazonie.
L’aire d’ O. paraguayensis , non loin de refuges définis par Haffer (1967) puis par Prance (1982), suggère une autre hypothèse. Le passage progressif en Bolivie de la forme « paraguayensis » à la forme boliviensis est typique d’un cline tel que défini par Huxley (1938), zoologiste. Ce concept, réactualisé par Salomon (2001, 2002), est maintenant admis par divers botanistes dont Raven et al. (2000).
| N |
Nanjing University |
| VI |
Mykotektet, National Veterinary Institute |
| J |
University of the Witwatersrand |
| E |
Royal Botanic Garden Edinburgh |
| V |
Royal British Columbia Museum - Herbarium |
| J |
University of the Witwatersrand |
| F |
Field Museum of Natural History, Botany Department |
| A |
Harvard University - Arnold Arboretum |
| AAU |
Addis Ababa University, Department of Biology |
| C |
University of Copenhagen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |