
Athanas ahyongi, Anker, Arthur & Komai, Tomoyuki, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.199308 |
|
DOI |
https://doi.org/10.5281/zenodo.6196514 |
|
persistent identifier |
https://treatment.plazi.org/id/03B687DE-FF92-8F55-FF74-A333FEC3FA70 |
|
treatment provided by |
Plazi |
|
scientific name |
Athanas ahyongi |
| status |
sp. nov. |
Athanas ahyongi View in CoL n. sp.
( Figs. 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Material examined. Holotype: female (CL 2.7 mm), Madagascar, Nosy-Bé, west of Hellville, 13°41.56’S, 48°24.65’E, sand flat with seagrass, rubble, patch reefs, some mounds of Glypturus sp., depth 0.5–1 m, suction (yabby) pump, from burrows or mounds, coll. A. Anker, 25 May 2008, fcn BMADA 0 466 ( FLMNH UF Arthropoda 14093). Paratype: male (CL 1.9 mm), same collection data as for holotype, fcn BMADA 0 467 ( FLMNH UF Arthropoda 14094). Non-type: 1 female (CL 2.6 mm), Japan, Ryukyu Archipelago, Okinawa Island, Red Beach, Kin, under rock, depth 8 m, coll. Y. Yamada, 4 March 2009 (CBM-ZC 10014).
Description (based on type specimens from Nosy-Bé). Small-sized alpheid shrimp. Body not particularly elongate, moderately slender. Carapace glabrous, not setose. Rostrum well developed, laterally compressed, horizontal, triangular in dorsal view, longer than broad; lateral margin slightly concave from rostral base to about mid-length, with some short setae; tip acute, not reaching distal margin of first segment of antennular peduncle; rostral carina faint, not reaching base of rostrum ( Fig. 1 View FIGURE 1 A, B). Extracorneal teeth well developed, triangular, sharp, not reaching anterior margin of cornea; infracorneal and supracorneal teeth absent ( Fig. 1 View FIGURE 1 A, B). Pterygostomial angle broadly rounded; cardiac notch deep ( Fig. 1 View FIGURE 1 A).
Pleura of first to fourth abdominal somites rounded posteroventrally; fifth pleuron with more pronounced posteroventral angle; sixth somite with large articulated triangular flap ( Fig. 1 View FIGURE 1 C). Telson widest at proximal third, distally tapering; dorsal surface with two pairs of strong spiniform setae inserted in deep pits; posterior margin slightly rounded medially, with two pairs of stout spiniform setae at each lateral angle, mesial almost twice as long as lateral; mesial spiniform setae obscured by long plumose setae originating from median area of posterior margin, between mesial spiniform setae ( Fig. 1 View FIGURE 1 D, E).
Eyes almost completely exposed in dorsal and lateral views; cornea large, normally pigmented ( Fig. 1 View FIGURE 1 A, B).
Antennular peduncle stout, with second segment squarish, about as long as wide, shorter than dorsally visible portion of first segment, equal to third segment; stylocerite sharp, reaching to about 0.7–0.8 length of second segment ( Fig. 1 View FIGURE 1 A); ventromesial carina with very strong, anteriorly produced tooth, as illustrated ( Fig. 1 View FIGURE 1 G); lateral flagellum biramous, fused portion composed of five segments. Antenna with basicerite stout, bearing small subacute tooth distoventrally; scaphocerite slightly exceeding end of antennular peduncle, ovate; blade broad, with convex anterior margin reaching beyond distolateral tooth; carpocerite distinctly exceeding scaphocerite ( Fig. 1 View FIGURE 1 A, B).
Mouthparts not dissected, appearing typical for genus in external view. Third maxilliped with coxa bearing large, distally acute lateral plate above mastigobranch; antepenultimate segment slender, somewhat flattened; penultimate segment almost four times as long as wide proximally, distally slightly widening; ultimate segment furnished with transverse rows of thick serrulate setae, tip without spiniform setae ( Fig. 2 View FIGURE 2 A).
Chelipeds symmetrical in shape, equal in size ( Fig. 3 View FIGURE 3 A, C), similar in both sexes, slightly more slender in male; ischium elongate, at least six times as long as wide, with ventrolateral margin strongly toothed, distal margin with three spiniform setae proximally; merus elongate, about as long as ischium, ventral surface, especially distally, flattened, ventrolateral margin strongly toothed; carpus vase-shaped, distally broadening, somewhat depressed laterally; chela laterally somewhat compressed, smooth except for areas of minute, barely visible granules on ventrolateral surface; fingers shorter than palm, feebly curved, with a moderately dense brush of elongate stiff setae originating from lateral surface of both fingers ( Fig. 3 View FIGURE 3 A); dactylus and pollex each with large triangular tooth fitting in a deep gap on opposable margin, tooth on pollex being more proximal than tooth on dactylus; pollex with additional, much smaller tooth adjacent and more distal to large tooth ( Fig. 3 View FIGURE 3 B, D).
Second pereiopod with ischium shorter than merus, both unarmed; carpus five-segmented, first segment longest, about the same length as remaining four segments, ratio of carpal segments (proximal to distal) approximately: 4: 1: 1: 1: 2; chela simple, longer than most-distal carpal article, with fingers slightly longer than palm ( Fig. 2 View FIGURE 2 B). Third pereiopod slender; ischium with one or two spiniform seta(e) on ventrolateral surface, dorsal margin with small spiniform seta proximally; merus more than five times as long as wide, unarmed; carpus conspicuously more slender than merus, about half as long as merus, distoventral angle with small spiniform seta; propodus as wide as carpus, ventral margin with three small spiniform setae, including a longer distal spiniform seta adjacent to dactylus; dactylus about 0.6 propodus length, simple, extremely slender, gradually curving distally ( Fig. 2 View FIGURE 2 C). Fourth pereiopod generally similar to third. Fifth pereiopod similar to third or fourth, but more slender; ischium with one small spiniform seta; propodus without spiniform setae, instead with three rows of thick serrulate setae on distolateral surface, most-distal row bearing longest setae ( Fig. 2 View FIGURE 2 D).
First pleopod with simple endopod, furnished with some setae, reaching half-length of exopod. Male second pleopod with appendix masculina shorter than appendix interna, apex with three slender setae ( Fig. 2 View FIGURE 2 E). Uropod with lateral lobe of protopod obscurely bifid, with small subacute point; diaeresis straight, ending in small acute tooth distolaterally, adjacent to slender distolateral spiniform seta ( Fig. 1 View FIGURE 1 F).
Gill-exopod formula typical for genus (see Anker & Jeng 2007); no rudimentary exopods visible on basis of P1–2.
Colour in life. Ground colour semitransparent greyish; most of mediodorsal area of carapace and abdomen occupied by broad white to pale yellowish band, extending from rostrum to sixth abdominal somite and continuing onto telson; flanks of carapace and abdomen with broad red band extending from anterolateral margin of carapace below eyes to sixth abdominal somite and continuing onto uropods; chelipeds and second to fifth pereiopods semitransparent-whitish; antennular peduncle partly colourless, partly white-yellowish, stylocerite bright red, antennular flagellum pale-orange; antennal basicerite, carpocerite and scaphocerite conspicuously red, antennal flagellum red proximally, colourless distally; corneas greyish-blue ( Fig. 6 View FIGURE 6 ).
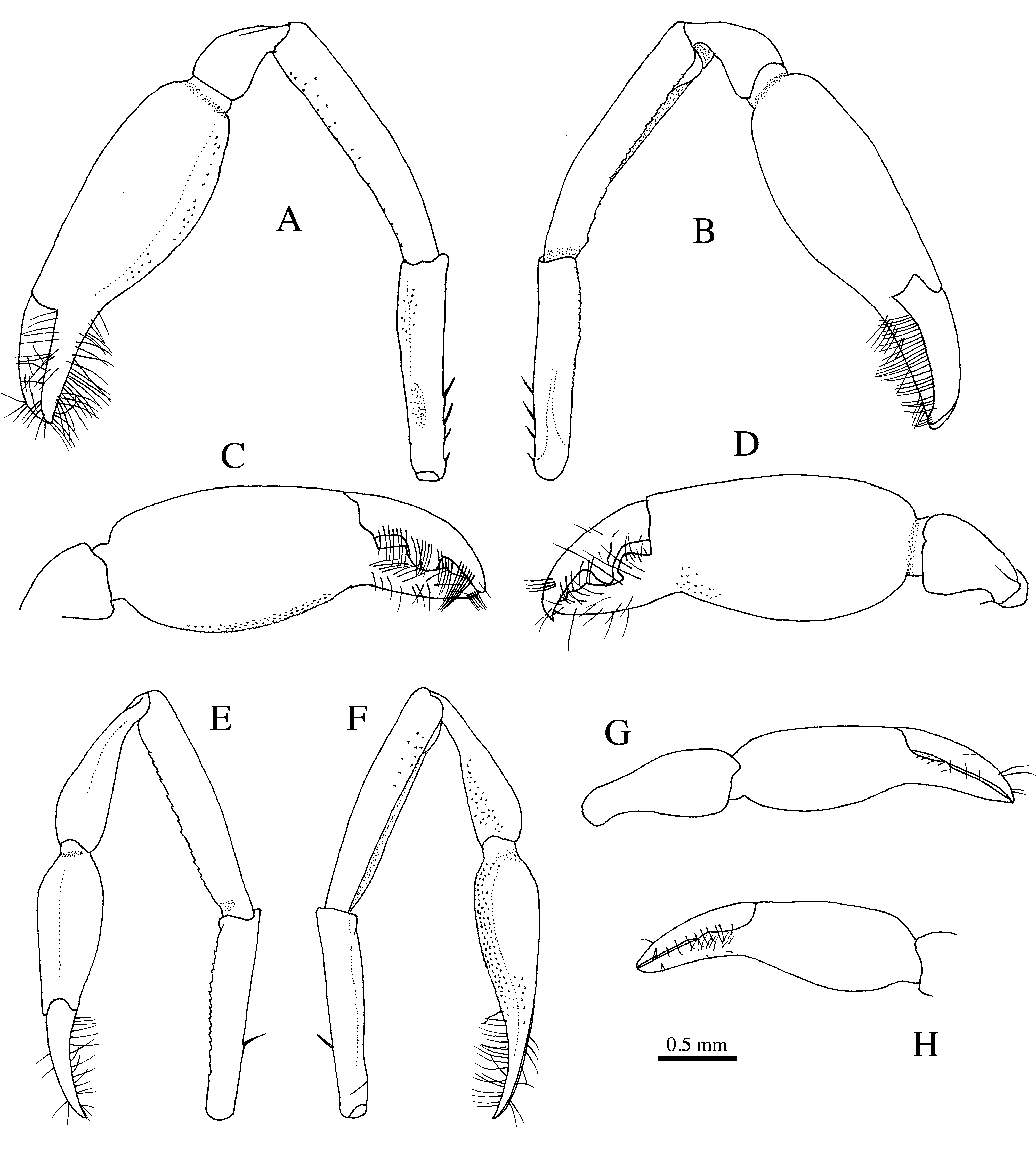
Variation. The non-type female from Okinawa differs from the two type specimens from Madagascar in several features, including the slightly shorter stylocerite, reaching to only half the length of the second segment of antennular peduncle ( Fig. 4 View FIGURE 4 A), and the degree of symmetry and proportions of the chelipeds ( Fig. 5 View FIGURE 5 ). While in the female holotype, the chelipeds are almost perfectly equal in size and symmetrical in shape, the chelipeds of the female from Okinawa are both unequal and asymmetrical, i.e. it has a major cheliped ( Fig. 5 View FIGURE 5 A-D) and a minor cheliped ( Fig. 5 View FIGURE 5 E-H). The major cheliped of the Okinawa female is generally similar to the cheliped of the female holotype, except for the much shorter carpus. The minor cheliped differs from the major cheliped by the distinctly longer carpus (longer than the carpus in the holotype), the much smaller chela (only about 30–40% of the volume of the large chela), and the absence of a strong armature on the fingers. These differences are here interpreted as intraspecific in view of the fact that the cheliped polymorphism is not uncommon in Athanas (e.g. Kemp 1915; Banner and Banner 1983; Anker 2003; Anker and Ahyong 2007).
Etymology. Named after our friend and colleague, Dr. Shane T. Ahyong (Australian Museum, Sydney) for his numerous contributions to decapod and stomatopod taxonomy and phylogeny.
Type locality. Nosy-Bé, Madagascar.
Distribution. Presently known only from northwestern Madagascar (Nosy-Bé) and Ryukyu Islands (Okinawa) in Japan; these two geographically remote records suggest that the species should be much more widespread in the Indo-West Pacific.
Ecology. The Nosy-Bé specimens were collected from burrows or mounds of an unknown host on a shallow seagrass-rubble flat, approximately 500 m from the shore, at a low tide depth of 0.5– 1 m. The Okinawa specimen was found on sand under a large rock, at a depth of 8 m, according to the collector’s field notes. However, turning over large rocks, especially those deeply embedded in the sediments, often exposes horizontal or oblique sub-surface burrows (A. Anker, pers. obs.); therefore, a possible association of this individual with a larger burrow cannot be excluded. The present evidence suggests that A. ahyongi sp. nov. is at least a facultative, burrow-associated commensal.
Remarks. Among the 28 species of Athanas present in the Indo-West Pacific ( Anker and Jeng 2007; Anker and Ahyong 2007; Anker and Marin 2007), only a few species share with A. ahyongi sp. nov. the combination of the following features: (1) the frontal margin of the carapace without infracorneal and supracorneal teeth; (2) the chelipeds more or less symmetrical in shape and subequal in size, with slender ischium and non-inflated merus and elongate, vase-shaped carpus; and (3) the third to fifth pereiopods with a simple, long, slender, sickle-shaped dactylus. These species are A. sydneyensis Anker and Ahyong, 2007 from New South Wales, Australia; A. squillophilus Hayashi, 2002 from Japan; and A. phyllocheles Banner and Banner, 1983 , a deep-water species from Réunion Island ( Banner and Banner 1983; Hayashi 2002; Anker and Ahyong 2007).
Athanas ahyongi sp. nov. differs from A. sydneyensis by the longer stylocerite, reaching to 0.7–0.8 length of the second segment of antennular peduncle (vs. reaching only to 0.5 in A. sydneyensis ); the proximolaterally non-convex, shorter rostrum, not reaching the distal margin of the first segment of antennular peduncle (vs. proximolaterally convex, reaching the distal margin in A. sydneyensis ); the stouter antennular peduncle, with the second segment as long as wide (vs. longer than wide in A. sydneyensis ); the much shorter cheliped carpus, which is only about 0.5 length of the merus (vs. almost equal to the merus in A.
sydneyensis View in CoL ); and the more gaping cheliped fingers, with a different configuration of teeth (see Anker and Ahyong 2007). Athanas ahyongi View in CoL sp. nov. can be easily separated from A. squillophilus View in CoL by the pterygostomial angle being rounded (vs. sharply produced in A. squillophilus View in CoL ); the cheliped ischium and merus having a dentate ventrolateral margin (vs. smooth in A. squillophilus View in CoL ); and the setal brush on the fingers of the chela not continuing onto the palm, as is the case in A. squillophilus View in CoL (see Hayashi 2002). The new species markedly differs from A. phyllocheles View in CoL by the general shape of the chelipeds and the much more slender antennular peduncle, second and third pereiopods (see Banner and Banner 1983).
All the other Indo-West Pacific species of Athanas View in CoL either have infracorneal and/or supracorneal teeth or differently shaped chelipeds, and sometimes both (e.g. Pearson 1905; De Man 1911, 1915, 1920; Chace 1955; Banner and Banner 1960, 1966, 1973; Bruce 1990; Anker et al. 2001; Hayashi 2002; Anker 2003; Anker and Marin 2007; Anker et al. 2010). The chelipeds of A. tenuipes De Man, 1910 View in CoL and A. gracilipes Banner and Banner, 1978 View in CoL are unknown, but these deep-water species differ from A. ahyongi View in CoL n. sp. by the presence of small blunt infra-corneal teeth and several other features. For instance A. tenuipes View in CoL differs by the longer and more slender rostrum, and A. gracilipes View in CoL by the much more slender second pereiopod and the distinctly shorter stylocerite ( De Man 1910, 1915; Banner and Banner 1978).
Among the Eastern Atlantic species, only A. amazone Holthuis, 1951 View in CoL and A. ivoiriensis Anker and Ahyong, 2007 View in CoL appear to be distantly related to the new species, from which they can be separated by numerous characters. Athanas ivoiriensis View in CoL differs by the much shorter antennal carpocerite, A. amazone View in CoL by the presence of a small infracorneal tooth, and both of them by the general shape and armature of the fingers of the chelipeds (cf. Holthuis 1951; Anker and Ahyong 2007). The new species also differs from all species of Athanas View in CoL with known colour patterns by its unique, longitudinally red-white striped colour pattern ( Fig. 6 View FIGURE 6 ), in particular the presence of a well-delineated, broad dorsolateral red band running from the antenna and anterior margin of the carapace to the sixth abdominal somite. Most other species of Athanas View in CoL have variously spotted or uniform reddish or purplish patterns, with or without a white mediodorsal band ( Anker & Jeng 2007), whereas two echinoderm-associated species have a red-white transversely banded pattern ( Marin et al. 2005; Anker and Marin 2007).
Athanas ahyongi View in CoL sp. nov. is the fifth known species of Athanas View in CoL found in association with burrows of larger animals. The other species are Athanas amazone View in CoL , found on a few occasions in burrows of the stomatopod Squilla mantis (L) in the Mediterranean Sea ( Froglia and Atkinson 1998); Athanas squillophilus View in CoL , associated with burrows of the stomatopod Oratosquilla oratoria (De Haan) in Japan ( Hayashi 2002); Athanas dentirostris Anker, Jeng and Chan, 2001 View in CoL , of which the single known specimen was found in a burrow of the mud-shrimp Austinogebia View in CoL aff. takaoensis (Sakai and Türkay) in Vietnam ( Anker et al. 2001); and Athanas japonicus Kubo View in CoL , occasionally associated with mudskippers, Periphthalmodon sp., in Australia ( Anker 2003) and other burrowing hosts (A. Anker, unpubl. data). The nature of these associations (facultative vs. obligate, mutualism vs. commensalism) as well as the hosts of A. ahyongi View in CoL sp. nov. remain to be determined.





| FLMNH |
Florida Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Athanas ahyongi
| Anker, Arthur & Komai, Tomoyuki 2010 |
A. ivoiriensis
| Anker and Ahyong 2007 |
Athanas dentirostris
| Anker, Jeng and Chan 2001 |
A. gracilipes
| Banner and Banner 1978 |
A. amazone
| Holthuis 1951 |
A. tenuipes
| De Man 1910 |