
Theodoxus fluviatilis, (LINNAEUS 1758)
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2003.00058.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B6B923-EE32-FFE5-8C2F-60D439C16ADE |
|
treatment provided by |
Carolina |
|
scientific name |
Theodoxus fluviatilis |
| status |
|
THEODOXUS FLUVIATILIS (LINNAEUS 1758) View in CoL
Material examined
Poitiens, France ( USNM 836154). Only preserved material was available for study. Consequently, scanning electron micrographs of complete midgut morphology were difficult to obtain and were supplemented with camera lucida drawings of dissected material. In Theodoxus fluviatilis , the midgut is rotated ~45∞ counter-clockwise within the visceral mass relative to caenogastropods (see Discussion). Thus, to facilitate topological comparisons with ingroup taxa, the midgut of was opened by a longitudinal incision along the left side.
External anatomy and mantle cavity
Anterior pedal gland opening anteriorly to shallow flap along broad propodium. Operculum present. Vestigial right gill present at base of mantle cavity below pallial glandular gonoduct.
Reproductive system
Gonopericardial canal absent. Proximal tip of glandular gonoduct embedded in visceral mass. Pallial portion of glandular gonoduct closed and diaulic. One genital opening leading to connected copulatory bursa and seminal receptacle; the former storing unorientated sperm, the latter orientated sperm.
Distal vas deferens forming highly coiled seminal vesicle. Prostate closed with distal genital aperture. Open seminal groove leading from genital aperture to penis lying on cephalic lappet to left of right cephalic tentacle.
Alimentary system
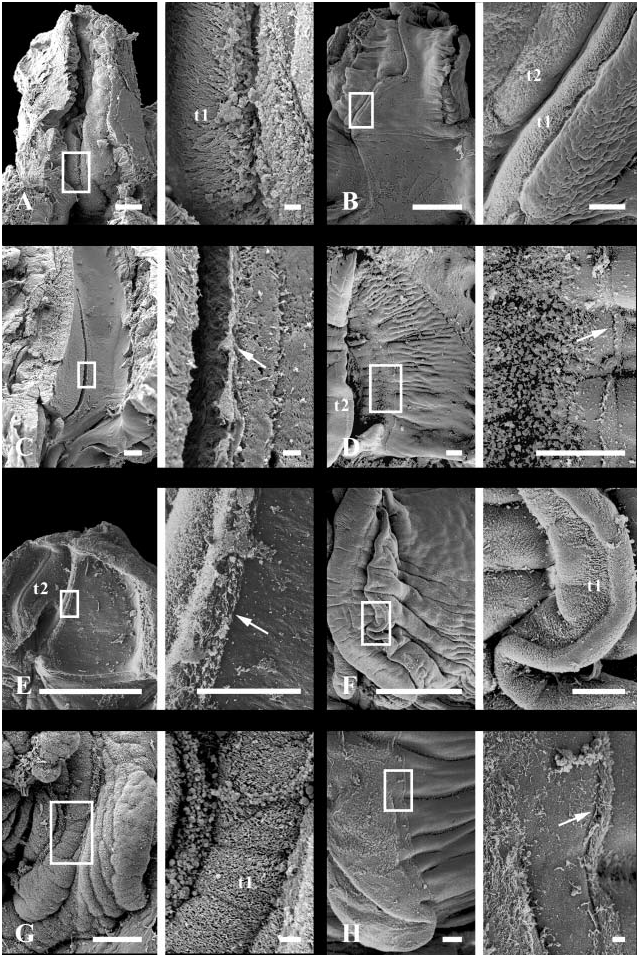
Foregut. Radula rhipidoglossate. Three paired odontophoral cartilages and one unpaired sublingual cartilage present. Subradular membrane completely covering odontophore to base of deep sublingual cavity; subradular organ absent ( Fig. 4A View Figure 4 ). Paired glands opening ventrally to sublingual cavity. Dorsal epithelium of buccal cavity cuticularized. Discrete jaw and salivary glands absent. Mid-ventral fold low in buccal cavity ( Fig. 8B View Figure 8 , vf), continuous with dorsal lip of radular sac. Paired lobes of glandular tissue developing below mid-ventral fold within anterior oesophagus. Low, weakly glandular ventro-lateral folds present along inner aspect of ducts to oesophageal pouches ( Fig. 9B View Figure 9 , vlf). Glandular mid-ventral fold persisting within anterior and mid-oesophagus, through region of torsion, before diminishing. Paired oesophageal pouches present, simple anteriorly, becoming increasingly folded and glandular posteriorly, comprising continuous buccal pouches ( Fig. 9B View Figure 9 , bp) and oesophageal glands ( Fig. 8B View Figure 8 , eg).
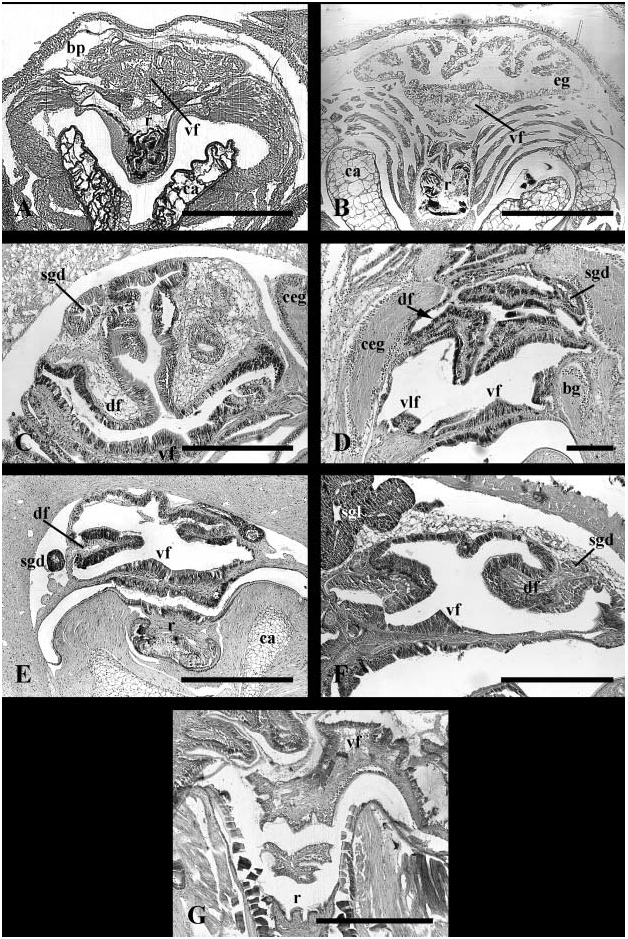
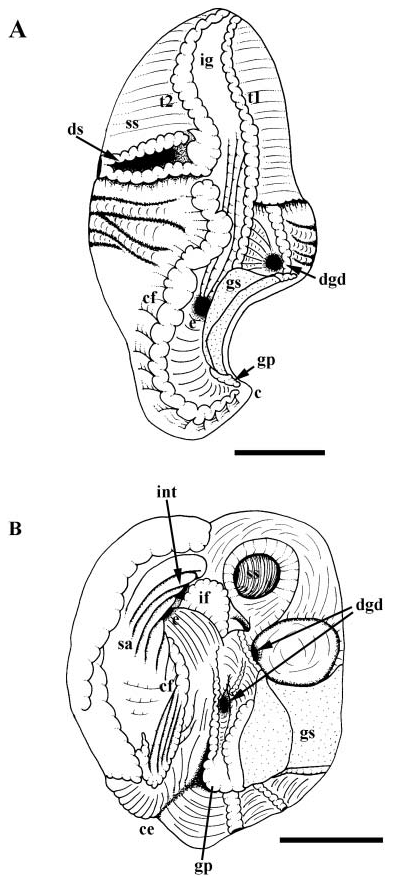
Midgut. Oesophageal aperture ( Fig. 12A, e View Figure 12 ) and posterior digestive gland duct lying to left of gastric shield (gs); anterior digestive gland duct (dgd) within deep pocket between gastric shield and style sac. Gastric shield dorsal, functionally separating oesophageal aperture and posterior digestive gland duct from gastric lumen. Ciliated fold (cf) beginning at proximal tip of minor typhlosole (t2), extending posteriorly to left of oesophagus. Ciliated fold and glandular pad (gp) curving into caecum (c) lying behind shield. Major typhlosole (t2) and several folds continuous with intestinal groove terminating at oesophageal aperture. Raised ciliary tract on major typhlosole absent ( Fig. 21A View Figure 21 ). Deep sac ( Fig. 12A View Figure 12 , ds) present, bounded posteriorly by lip of style sac and anteriorly by second fold, both continuous with minor typhlosole. Mouth of sac surrounded by differentiated style sac cilia. Crystalline style absent.
Hindgut. Intestine completing approximately two loops. Rectum penetrating ventricle.
Reno-pericardial system
Transverse pallial vein extending dorsally from cephalopedal sinus, splitting into afferent branchial vein and afferent renal vein. Afferent branchial vein traversing anterior wall of kidney to afferent ctenidial membrane; afferent renal vein passing through thinwalled ventral chamber of kidney (= bladder), supplying dorsal chamber containing mass of excretory lamellae. Vestigial right gill present. Blood sinus of vestigial right gill communicating directly with cephalopedal sinus. Nephridial gland absent.
Nervous system and sensory structures Circum-oesophageal nerve ring surrounding buccal mass anteriorly, with buccal connectives passing posteriorly to buccal ganglia. Nervous system hypoathroid, right zygoneurous and left dialyneurous, with paired visceral ganglia. Labial commissure present. Single pedal commissure connecting pedal ganglia. Pedal cords present. Tentacular nerve single. Statocysts with single statolith present dorsally and medially, just behind pedal commissure. Eyes on tips of short ocular peduncles.
Remarks
This species has been the subject of many anatomical investigations, emphasizing the mantle cavity, reproductive and alimentary systems. Gilson (1896), Fretter & Graham (1962) and Fretter (1946) provided information for reproductive anatomy; the latter account is the most comprehensive and details gross morphology as well as histological fine structure for both males and females.
Lenssen (1899), Bourne (1908), Graham (1939) and Whitaker (1951) provided information for the foregut. Whitaker’s account, describing the structure and function of the foregut, is the first to suggest that the outpocketings of the foregut represent continuous buccal pouches and oesophageal glands; these structures had been variously interpreted as salivary glands ( Lenssen, 1899) or simply as oesophageal pouches ( Bourne, 1908; Graham, 1939). Whitaker was also the first to recognize the presence of paired sublingual glands in this species. Whitaker acknowledges the presence of the mid-ventral fold, but does not describe the presence of ventro-lateral folds. The species possesses a seventh unpaired cartilage ( Lenssen, 1899; Bourne, 1908; Fretter, 1946); however, this supports the cuticle of the sublingual pouch and is not considered to represent part of the odontophore (cf. Sasaki, 1998).
Preliminary observations on midgut and hindgut anatomy were provided by Lenssen (1899) and were described more completely by Graham (1939) and Fretter & Graham (1962); Graham clarified the absence of the crystalline style in this species. Several findings obtained here for midgut morphology differ subtly from those reported previously ( Graham, 1939; Fretter & Graham, 1962). In the present analysis, the oesophageal aperture and one digestive gland duct were found immediately to the left of the gastric shield such that, when the midgut is closed, the shield separates them from the gastric chamber. A second digestive gland duct is present within the deep pocket between the gastric shield and style sac.
Landsburg (1882) and Lenssen (1902) provided the main points of reno-pericardial morphology and circulation, which were elaborated by Bourne (1908); Fretter (1965) primarily described blood flow through the mantle and associated organs. Blood supply to the vestigial right gill, an organ first noted by Lenssen (1902), has been attributed to a diverticulum off the transverse pallial vein ( Bourne, 1908; Fretter, 1965). However, as described above, histological sections revealed that the vestigial gill sinus opens directly to the cephalopedal sinus.
Lenssen (1902) provided a relatively thorough account of the nervous system which was refined by Bourne (1908).
Discussion
Reproductive anatomy of most neritids is highly complex, particularly among females. The pallial oviduct is typically diaulic, one aperture leading to the bursa and seminal receptacle, the second leading to the glandular oviduct, which is posteriorly embedded in the visceral mass and subdivided into capsule and albumen glands ( Bourne, 1908; Fretter, 1946, 1965; Starmühlner, 1969; Berry, Lim & Sasekumar, 1973; Houston, 1990; Haynes, 1991; Sasaki, 1998). These two parallel portions of the oviduct communicate posteriorly via a narrow duct. Some neritids possess a triaulic pallial oviduct. The third aperture is referred to as the ductus enigmaticus ( Bourne, 1908) and, as the name implies, is of uncertain function but has been suggested to regulate internal pressure by allowing water into the oviduct or excess sperm to escape ( Bourne, 1908; Fretter, 1965; Houston, 1990; Haynes, 1991). In addition, some neritids have a small duct that leads from the renal oviduct and opens to the posterior end of the mantle cavity adjacent to the vestigial right gill ( Fretter, 1965). The presence of this duct, its position within the body and its positional relationship to the vestigial gill has been interpreted as evidence that the right kidney has been integrated into the glandular reproductive tract ( Fretter, 1965), rather than forming the seminal receptacle, as suggested by Thiele (1902). This connection between gonoduct and mantle cavity is absent in Theodoxus fluviatilis ( Fretter, 1946) . Bourne (1908) described the so-called oviduco-coelomic funnel (= gonopericardial canal), which connects the gonoduct with the extensive pericardial coelom. Despite careful searching, Fretter (1965) was unable to confirm this.
Male reproductive anatomy typically consists of a testis, a distal vas deferens that is highly coiled and functions in the storage of sperm, and a highly glandular prostate that forms the spermatophore. The prostate is closed, discharging through an aperture adjacent to the anus, and is subdivided into two distinct masses of glands. The penis, when present, lies on the cephalic lappet, to the left of the right cephalic tentacle ( Bourne, 1908; Andrews, 1937; Starmühlner, 1969; Houston, 1990; Sasaki, 1998).
The relatively conservative foregut anatomy consists of paired sublingual glands opening ventrally to the buccal cavity, and confluent buccal pouches and oesophageal glands lying anterior to the region of torsion; salivary glands are absent ( Thiele, 1902; Bourne, 1908; Whitaker, 1951; Fretter, 1965). There are typically three sets of paired cartilages ( Lenssen, 1899; Bourne, 1908; Fretter, 1946; Salvini-Plawen, 1988); the median cartilages may be fused or unfused and a seventh unpaired cartilage may be present below the anterior tip of the subradular membrane ( Sasaki, 1998).
In general, the neritid midgut consists of a globular gastric chamber that bears a highly variable number of digestive gland duct openings; a small, textured pouch posterior to the gastric shield may be present ( Haller, 1894; Bourne, 1908; Fretter & Graham, 1962; Fretter, 1965). A prominent fold associated with the minor typhlosole passes posteriorly into the caecum; this fold may be absent or elaborated into a well-developed sorting area ( Fretter, 1965). Theodoxus fluviatilis remains the only neritid described possessing a small transverse pouch just within the lip of the style sac ( Graham, 1939). However, a similar pouch has been observed in Neritina reclivata (pers. obs.).
The neritid reno-pericardial system comprises an enlarged pericardial coelom, a heart with two auricles, and a long, ciliated reno-pericardial canal that opens to the dorsal, glandular portion of the two-tiered kidney ( Fretter, 1965). The ventral portion of the kidney forms a bladder into which the nephropore opens ( Lenssen, 1902; Thiele, 1902; Bourne, 1908; Sasaki, 1998; Estabrooks, Kay & McCarthy, 1999). The intestine pierces not only the ventricle, but the lumen of the kidney as well ( Thiele, 1902), posteriorly separating a portion of the reno-pericardial coelom.
The neritid nervous system is right zygoneurous and left dialyneurous, with paired visceral ganglia – one lying in the transverse pallial vein and the second adjacent to the vestigial gill ( Bouvier, 1892; Bourne, 1908). The pedal ganglia are united by a single commissure ( Bourne, 1908; Sasaki, 1998). The identity and homology of the subintestinal ganglion, while previously in question ( Haller, 1894), consists of a swelling lying between and just behind the pleural ganglia ( Bouvier, 1892; Bourne, 1908; Starmühlner, 1969).


| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |