
Chrysosphaerella rotundata Škaloudová & Škaloud, 2013
|
publication ID |
https://doi.org/10.11646/phytotaxa.130.1.4 |
|
DOI |
https://doi.org/10.5281/zenodo.5085775 |
|
persistent identifier |
https://treatment.plazi.org/id/03B787CE-5750-6303-FF46-65F0F620F849 |
|
treatment provided by |
Felipe (2021-07-09 14:18:37, last updated by Plazi 2023-11-03 07:30:16) |
|
scientific name |
Chrysosphaerella rotundata Škaloudová & Škaloud |
| status |
sp. nov. |
Chrysosphaerella rotundata Škaloudová & Škaloud , sp. nov. ( Figs. 1–6 View FIGURES 1–6 )
Colonies spherical, consisting of cells bearing two flagella. Cells covered by numerous scales and spines. Three size classes of scales occur: large circular scales (3.0–3.5 × 2.2–3.1 µm), large oval scales (2.0–3.1 × 1.3–2.4 µm), and smaller oval scales (1.5–1.7 × 1.2 µm). Scales patterned with a series of short ridges forming a scalloped shaped pattern. Spines with a shaft joining the two plates of the bobbin-like structure. The length of the spines varies from 4 µm to 10 µm. Cysts unknown.
Type:— FINLAND. Keski-Suomi, 62° 15’ 1.07” N, 26° 34’ 48.00” E, a small, oligotrophic unnamed lake, water temperature 5.6 °C, pH 7, conductivity 40 µS cm -1, coll. Škaloudová & Škaloud, 1 May 2012 (holotype: Strain S 89.C4, frozen material deposited at the Culture Collection of Algae of the Charles University in Prague, Department of Botany , Benátská 2, 12801 Prague 2, Czech Republic ). Fig. 2 View FIGURES 1–6 is an illustration of the holotype GoogleMaps .
Cells were grouped into spherical colonies. Individual cells were spherical to pyriform, about 12–13 Μm long and 11 Μm wide, bearing two flagella. Cells were covered by numerous scales and spines. Three size classes of scales might be discerned; however, a continuous transition in size of scales existed. The majority of the scales were larger and oval in outline ( Figs. 1, 3 View FIGURES 1–6 ). The second type of the scales, larger and circular in outline, were produced less frequently ( Fig. 2 View FIGURES 1–6 ). However, presence of the large, circular-shaped scales was a main distinguishing character of Chrysosphaerella rotundata . Both large circular and oval scales were patterned with a series of short ridges forming a scalloped shaped pattern. The pattern of smaller oval scales ( Fig. 3 View FIGURES 1–6 ) was the same as that of larger scales, but often less distinct. The spines had a thick shaft joining the two plates of the bobbin-like structure ( Figs. 4–5 View FIGURES 1–6 ); however, a variation in the thickness of the shaft was observed and spines with a thinner shaft were found as well ( Fig. 6 View FIGURES 1–6 ).
Etymology:— The specific epithet ‘rotundata’ refers to the rounded shape of scales.
Phylogenetic analyses, ITS2 secondary structures:— Bayesian inference, Maximum Likelihood, and Maximum Parsimony analyses inferred from the concatenated SSU rDNA and rbc L sequences resulted in highly similar phylogenetic trees, recognizing the six main lineages within Chrysophyceae ( Fig. 7 View FIGURE 7 ). According to their members, the lineages could be recognized as traditionally defined orders: Chromulinales , Chrysosaccales, Hibberdiales, Hydrurales, Ochromonadales , and Synurales . With the exception of the Ochromonadales , all lineages were also significantly supported by ML and wMP analyses. Both analyses resolved the Ochromonadales as monophyletic, but without any statistical support. The genus Chrysosphaerella formed a firmly supported monophyletic lineage within the Chromulinales ( Fig. 7 View FIGURE 7 ). It was divided into two supported subclades. Chrysosphaerella rotundata was inferred as closely related to morphologically similar C. brevispina , with which it formed the first inferred subclade. The species differed by eight and eleven nucleotide substitution changes in the SSU rDNA and rbc L sequences, respectively. The second subclade consisted of C. longispina and two environmental Chrysosphaerella isolates. Since the isolates were molecularly characterized by only the SSU rDNA (summer isolate) or rbc L (winter isolate) sequence, we could not exclude that they in fact belong to the same genotype. The environmental isolates differed by two nucleotide substitution changes in the SSU rDNA and rbc L sequences from the C. longispina sequences.
To further evaluate the degree of genetic differentiation of the closely related C. rotundata and C. brevispina , we additionally sequenced the ITS1- 5.8S rDNA-ITS2 region. The ITS1 region was hardly alignable, and the sequences differed significantly in their length (342 bp in C. rotundata vs. 294 bp in C. brevispina ). Within the aligned regions, the divergence between the ITS1 rDNA sequences was approximately 15.5%. The length of the ITS2 region was similar in both species (293 bp in C. rotundata vs. 287 bp in C. brevispina ), and the overall divergence between the ITS2 rDNA sequences was approximately 12.5%. In addition, we mapped the differences in the ITS2 sequence on a predicted ITS2 secondary structure ( Fig. 8 View FIGURE 8 ). Most of the differences were located in terminal and internal hairpin loops. Four compensatory base changes (CBCs) were found in the stem regions of helices I and II.
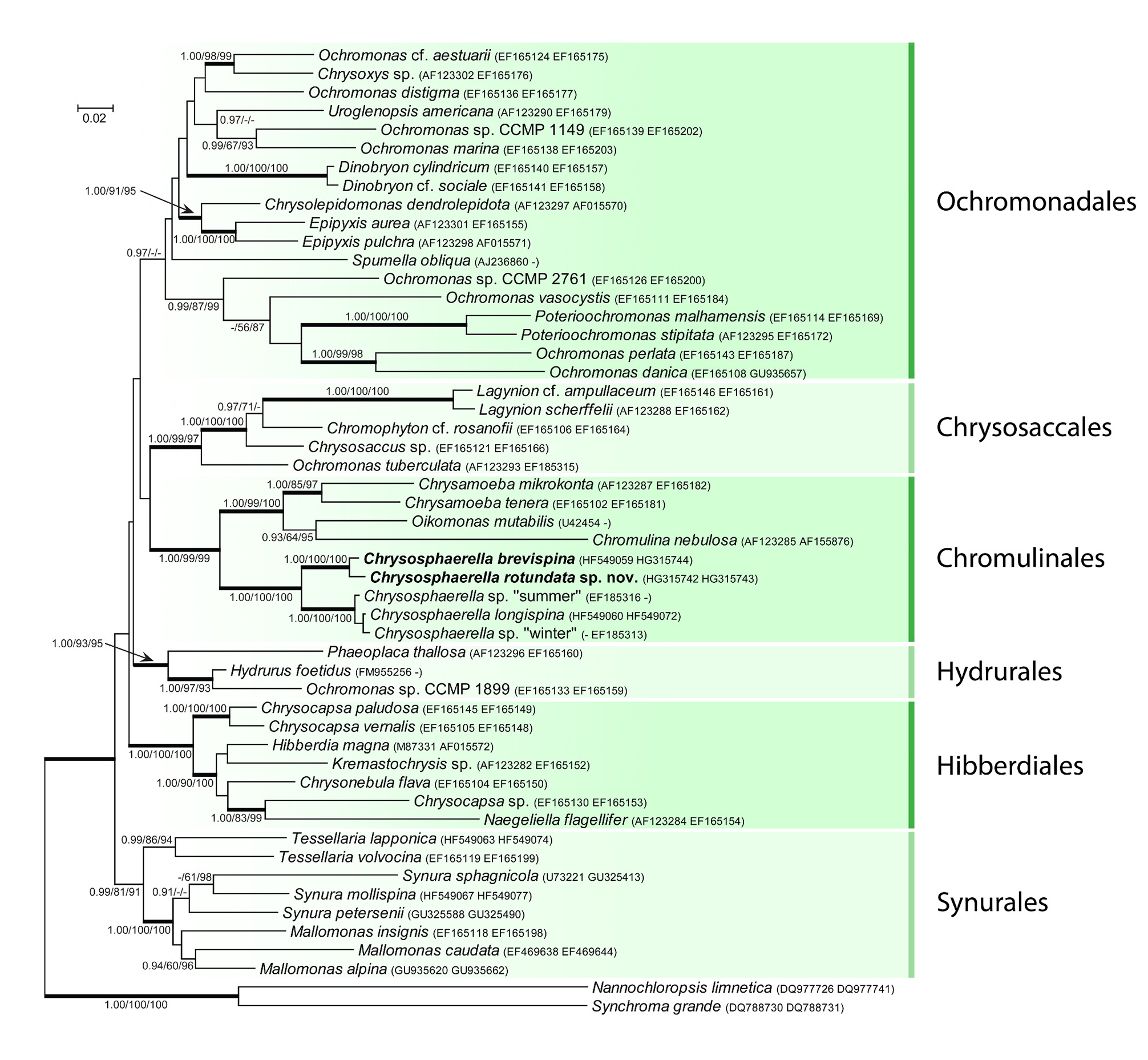
FIGURE 7. Bayesian analysis of Chrysophyceae, based on the combined and partitioned SSU rDNA + rbcL dataset using a GTR+G+I model for all partitions. Values at the nodes indicate statistical support estimated by three methods – MrBayes posterior-node probability (left), maximum-likelihood bootstrap (middle), and maximum parsimony bootstrap (right). Thick branches represent nodes receiving the highest PP support (1.00). Newly obtained sequences are given in bold. Accession numbers for the concatenated sequences (SSU rDNA and rbcL, respectively) accompany each species name. Scale bar shows the estimated number of substitutions per site.
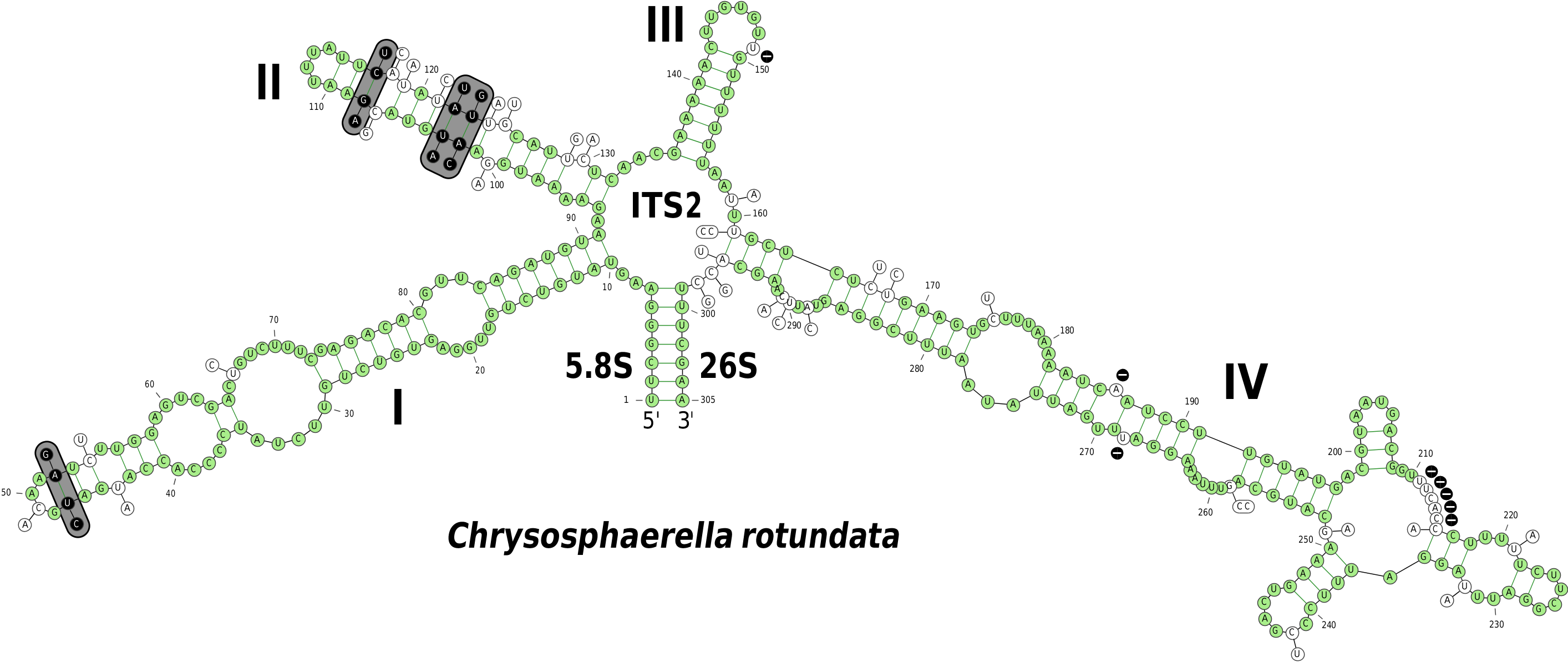
FIGURE 8. Comparison of the ITS2 sequences and predicted secondary structures of Chrysosphaerella rotundata and C. brevispina. Base numbering is indicated every 10 bases, and the four helices are numbered with Roman numerals. The structure shown corresponds to C. rotundata; positions conserved in C. brevispina are portrayed in green, bases substituted in C. brevispina are shown by the structure and connected to the respective position by a short line, deletions are indicated with minus symbols. Four base pairs marked in grey boxes are compensatory base changes (CBCs).
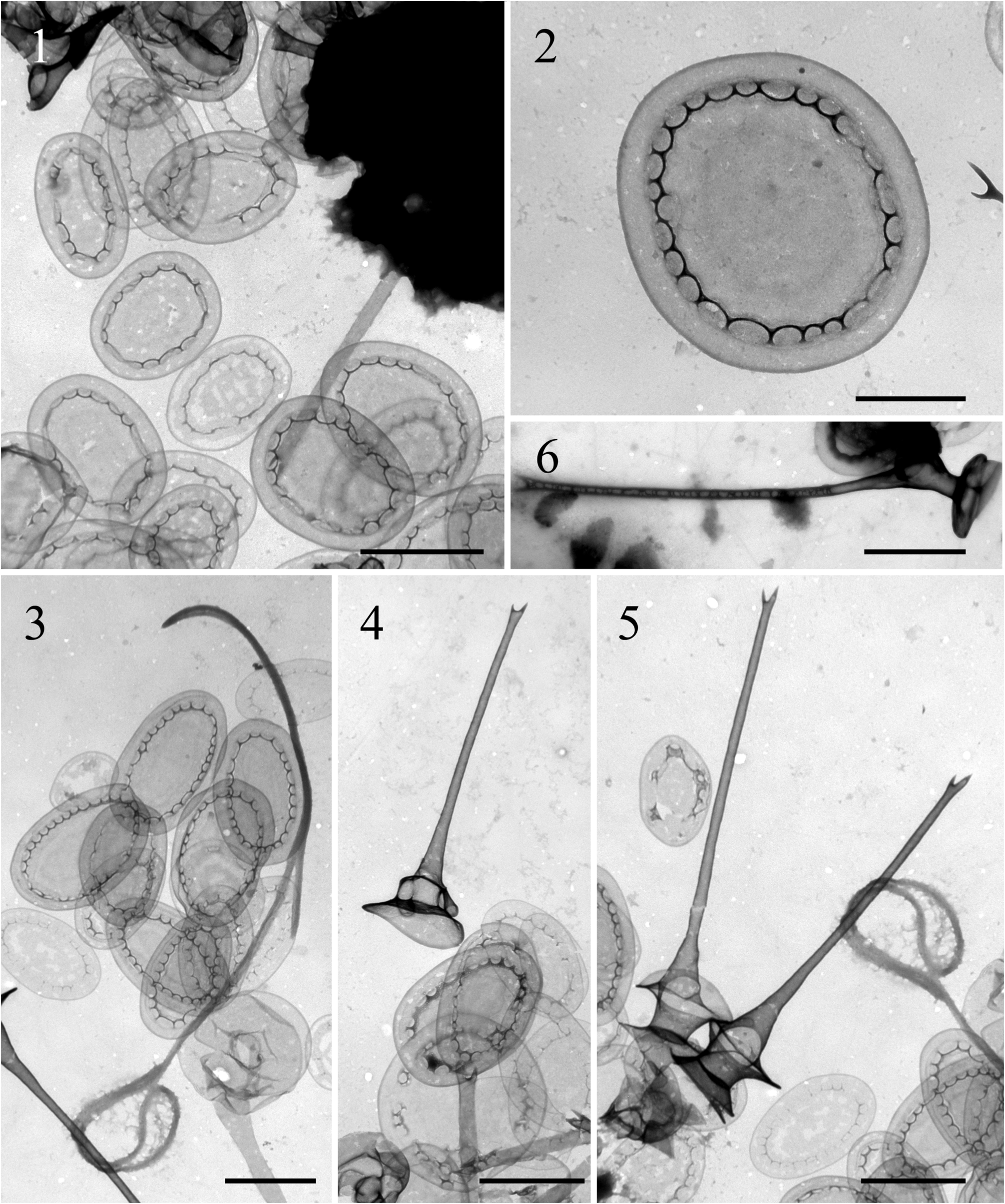
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |