
Atopocrinidae Messing (in Hess & Messing, 2011 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4731.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:85C4B527-6CAF-404B-87B9-B9FD5468213F |
|
DOI |
https://doi.org/10.5281/zenodo.3664749 |
|
persistent identifier |
https://treatment.plazi.org/id/03B89821-FF9C-E91F-7BAD-DF35FBD22925 |
|
treatment provided by |
Plazi |
|
scientific name |
Atopocrinidae Messing (in Hess & Messing, 2011 ) |
| status |
|
Family Atopocrinidae Messing (in Hess & Messing, 2011) View in CoL
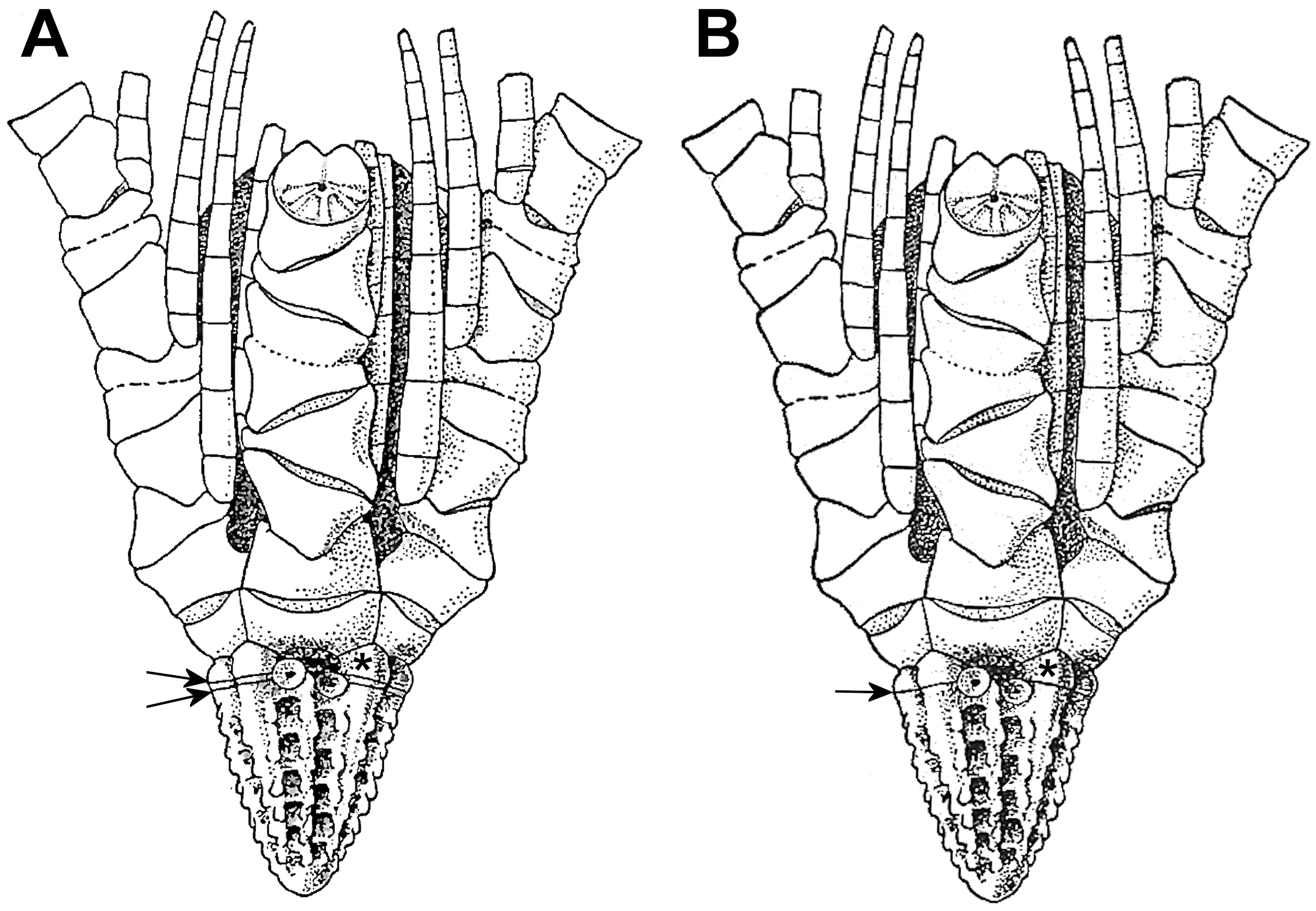
Diagnosis. Centrodorsal conical, as tall as or taller than wide, with narrow centrodorsal cavity and strong narrow interradial ridges usually crossed by one or more faint, superficial, suture-like lines adjacent to basal/centrodorsal suture; adoral surface with 5 interradial grooves that accommodate flattened, tongue-like basals. Cirrus sockets in 10 or 15 columns; each socket initially arising as a tube along adoral surface of centrodorsal; fully developed sockets with projecting, lateral, fulcral processes, convex or rounded-triangular in profile. Basals visible externally as triangular or pentagonal adoral extensions of centrodorsal interradial ridges. Arms 5, undivided. Synarthry between brachials 1 and 2. First syzygy between brachials 4 and 5. Articular facets of proximal three syzygial pairs (br4+5, 7+8 and 14+15) with only five radiating ridges. Proximal few pinnules arising from lateral projections of brachials; first pinnule on brachial 2. Pinnulars hollow, thin-walled (at least in A. sibogae ), and longer than wide, except short first two pinnulars on P3 and following pinnules; distal pinnulars unknown (modified from Hess & Messing 2011).
Type genus. Atopocrinus A.H. Clark, 1912 .
Remarks. Atopocrinidae includes only Atopocrinus , which is here construed to include two species. Although several characters are found in other extant crinoids, e.g., undivided rays (several families), first syzygy at br4+5 ( Pentametrocrinidae ), and cirrus sockets with strongly developed triangular fulcral ridges ( Atelecrinidae ), at least three characters, i.e., hollow pinnulars, suture-like lines crossing interradial ridges of centrodorsal, and proximal syzygies with only five radiating ridges, are unique. Additional characters may also prove unique (e.g., interradial buttresses in the centrodorsal cavity crossed by fine ridges and grooves) but have not been examined in the single known specimen of A. sibogae .
A.H. Clark (1912) placed Atopocrinus in Atelecrinidae with Atelecrinus but noted no characters in common except that its brachials resembled those of Atelecrinus . He later (A.H. Clark 1918) based its inclusion on the similar structure and arrangement of cirrus sockets, structure of the disk, and triangular proximal brachials and first syzygial pair in the two genera, which he considered different than in all other feather stars.
A.H. Clark (in A. H. Clark & A. M. Clark 1967) maintained Atopocrinus in Atelecrinidae and added Sibogacrinus (for Atelecrinus anomalus A.H. Clark, 1912 ), although the resulting familial diagnosis (A.M. Clark, in A. H. Clark & A. M. Clark 1967, p. 811) included no features common to all three genera, e.g., cirrus sockets flanked by “prominent elevations” sometimes forming a horseshoe-shaped ridge, and triangular proximal brachials (both absent in Sibogacrinus ), and proximal pinnules absent (except in Atopocrinus ). Rasmussen & Sieverts-Doreck (1978) added Upper Cretaceous-Lower Paleocene Jaekelometra , but their familial diagnosis likewise included numerous exceptions and only one possible feature common and unique to all four genera among extant feather stars: basal ossicles forming a ring of plates. However, these differed substantially among the genera (see below). They placed Atelecrinidae in the otherwise extinct (Lower Jurassic-Lower Paleocene) superfamily Paracomatulacea, another taxon with no synapomorphies. Gislén (1924) had placed Jaekelometra in the extinct family Conometridae and noted features that distinguished it from Atelecrinidae , e.g., much smaller centrodorsal cavity and very different radial articular faces. However, he diagnosed Conometridae chiefly on a combination of absent (e.g., no dorsal star) and widely variable characters (e.g., basals large and contiguous to hidden), with only one feature in common to all included genera: muscle fossae large in comparison with interarticular ligament fossae. However, this feature appears in several other unrelated feather star families, e.g., some Charitometridae , Zenometridae , and Pentametrocrinidae (A.H. Clark 1921) .
Messing (2003) removed Jaekelometra and Sibogacrinus from Atelecrinidae , based chiefly on basal ossicle features. However, Hess & Messing (2011) returned the latter to Atelecrinidae (in its own superfamily Atelecrinoidea), based on a re-examination of the type of Sibogacrinus , but removed both Jaekelometra and Atopocrinus to their own families (Jaekelometridae and Atopocrinidae ) without superfamilial assignment. The removal of the latter was based on a preliminary examination of the specimen collected off Chiba, Japan, and described below. The family remains monogeneric and unassigned to superfamily.
Because basal ossicles have played a pivotal role in descriptions of Atopocrinus and Atelecrinidae , their treatment in feather stars is reviewed here. Note that, with the inclusion of stalked bourgueticrine and guillecrine crinoids in order Comatulida ( Rouse et al. 2013; Hemery et al. 2013), the terms feather star and comatulid (applied to the ordinal level) are no longer synonymous. Feather star is thus an informal term for those Comatulida that shed the postlarval stalk and take up a free existence.
Basal ossicles form a complete circlet in most fossil feather stars, including the oldest true Comatulida such as Lower Jurassic Palaeocomaster ( Hess & Messing 2011) , and in the larvae and postlarvae of extant feather stars. However, in extant feather stars that have been examined previously, except Atelecrinidae and apparently some Zenometridae (A.H. Clark 1915; Messing & White 2001), they supposedly transform from a complete circlet into an internal rosette that sits atop the centrodorsal cavity, with or without projecting structures called basal rays. The single specimen of Atopocrinus ojii n. sp. dissociated for scanning electron microscopy and described herein also lacks a rosette, but its presence or absence cannot yet be determined in A. sibogae .
The status of basal ossicles in Atopocrinus is unclear due to ambiguity about the nature of basals relative to basal rays. A.H. Clark (1915) treated basal rays as “Prismatic calcareous rods of secondary origin developed in the basal grooves between the radial pentagon and the centrodorsal” (p. 70) with “no connection with the larval basals” (p. 72), which metamorphose following the stalked pentacrinoid postlarval stage into an internal rosette. He cited P. H. Carpenter (1879), who described the development of basal rays in the feather star Actinometra (now Comatula spp. and Comanthus parvicirrus ; family Comatulidae ) as calcification of tissue lying in the interradial furrow between the centrodorsal and radials. However, Gislén (1924) treated both complete circlets and narrower rods as components of basal ossicles. Breimer (1978, p. T27) noted that “interradial processes of the rosette may connect with five rod-like structures, known as basal rays”, but he did not specify their independent origin. Moore et al. (1978, p. T233) also defined basal rays separately from basals. In contrast, Rasmussen & Sieverts-Doreck (1978, p. T868) treated basal rays without explanation as modified basal ossicles rather than separate entities: “In all other comatulids [excepting Atelecrinidae , Decameridae , and the oldest, such as Archaeometra ] the basals are more or less reduced. Generally, they form five slender, interradial rods lodging in the shallow interradial furrows in the centrodorsal.” Despite their differences, the latter three references all appeared in the same volume: Part T Echinodermata 2 of the Treatise on Invertebrate Paleontology.
The ambiguity is particularly acute with reference to Atopocrinus , because A.H. Clark (1912, p. 150) first wrote that “The five basals are extremely reduced so that they appear externally as five narrow lines separating the basal rays from the centrodorsal; the basal rays are large and well developed”, and later (1915, p. 329) that “Large basal rays occur, just proximal to the extremely reduced and laminar basals.” He also described the basals as unmetamorphosed (1915, pp. 318, 348). His figure (A.H. Clark 1915, fig. 227, p. 245), shows what he interpreted as the narrow basals and large pentagonal knoblike ends of basal rays ( Figure 1A View FIGURE 1 ). However, he afterwards (A.H. Clark 1918, pp. viii, 263; A.H. Clark in A.H. Clark & A.M. Clark 1967, p. 815) diagnosed the genus as having no basals but only that “High and narrow basal rays, of which the outer ends are broadly pentagonal and convex, cap the proximal ends of the interradial ridges and extend inward under the radial pentagon, forming the sides and the blind inner wall of the subradial clefts” ( Figure 1B View FIGURE 1 ). The illustration (A.H. Clark 1918, fig. 15, p. 263) is the same as that published in 1915, except that the line marking the aboral suture of the supposed “reduced and laminar basals” has been omitted. I have not found that he ever addressed the change or mentioned what might have been misinterpreted as the aboral line. The photograph of the specimen (A.H. Clark 1918, pl. 26, fig. 95) is too small to identify any of these features.
Whether or not the narrow basal rays characteristic of many Comatulidae and several other feather star taxa (e.g., Thalassometridae , Charitometridae , Calometridae , Ptilometridae , Heliometrinae) derive directly from embryonic basals or are secondary deposits, it seems less likely that the broad, tongue-like basal rays illustrated by Gislén (1924) for a young Asterometra anthus (Asterometridae) , and also found in some Zenometridae ( Messing & White 2001) , represent such secondary deposits. As a result, because the corresponding ossicles in Atopocrinus ojii n. sp. are similarly large and tongue-shaped, and no rosette appears to exist, they are treated here as basal ossicles rather than either secondary structures or basal rays. References to A.H. Clark’s initial distinction between basal rays and reduced basals, when needed, are included parenthetically to avoid confusion.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.