
Cataphractomimus impervius, Gustafsson & Lei & Chu & Zou, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4742.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:CA0AD801-C329-4D41-B081-1647491DF842 |
|
DOI |
https://doi.org/10.5281/zenodo.3684875 |
|
persistent identifier |
https://treatment.plazi.org/id/03BA7024-9B1B-EC5E-55EF-FDD0FB31F822 |
|
treatment provided by |
Plazi |
|
scientific name |
Cataphractomimus impervius |
| status |
sp. nov. |
Cataphractomimus impervius new species
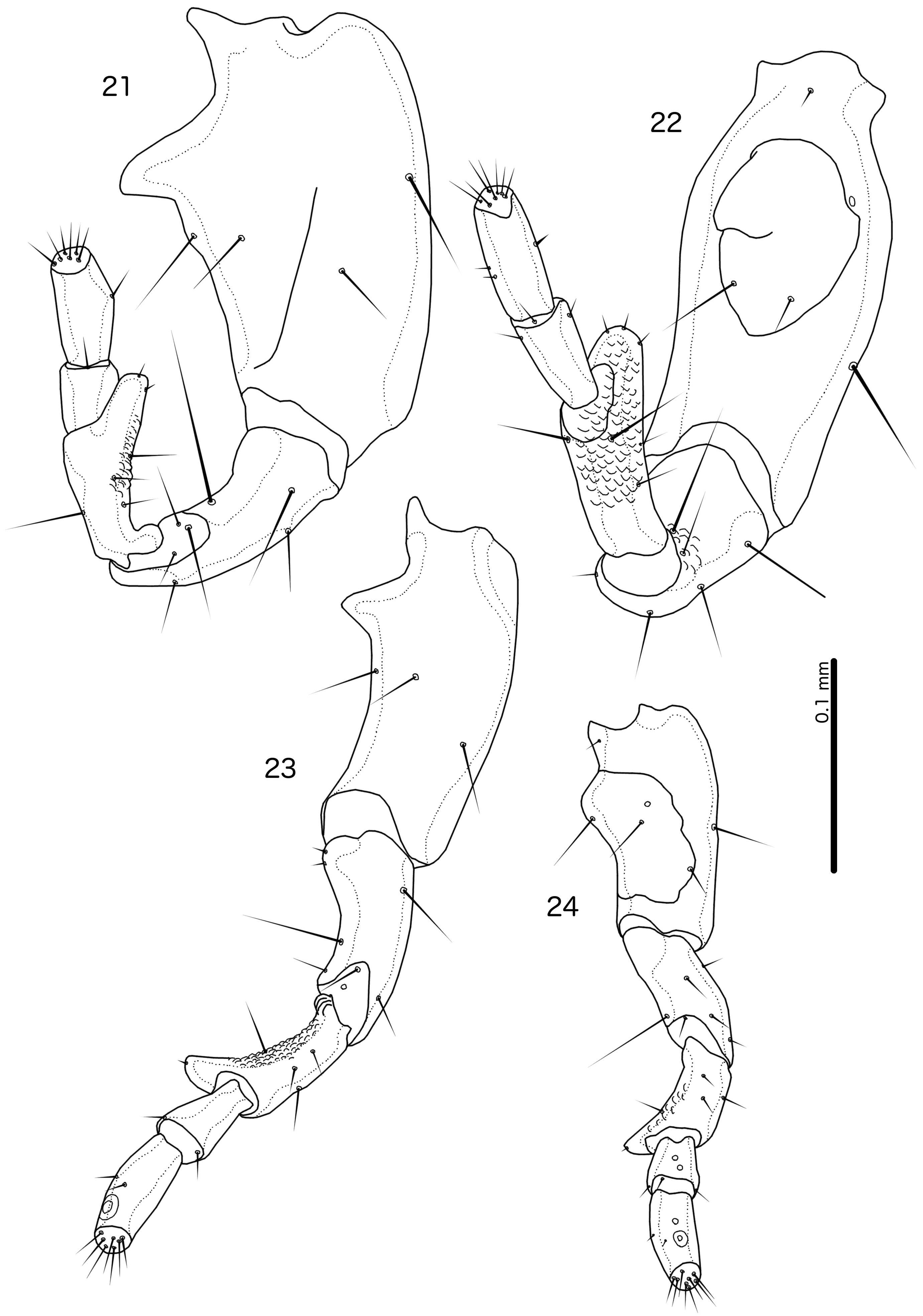
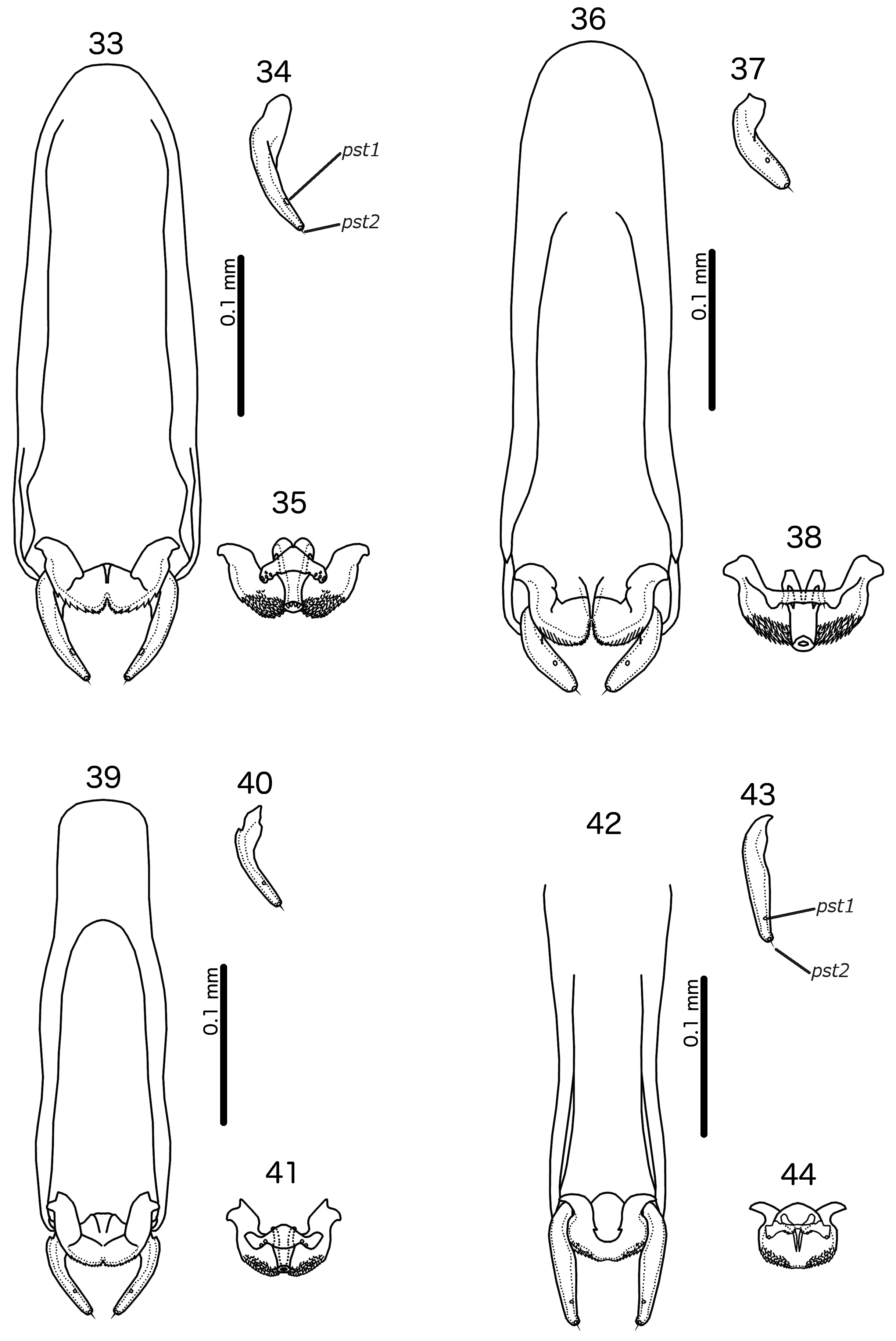
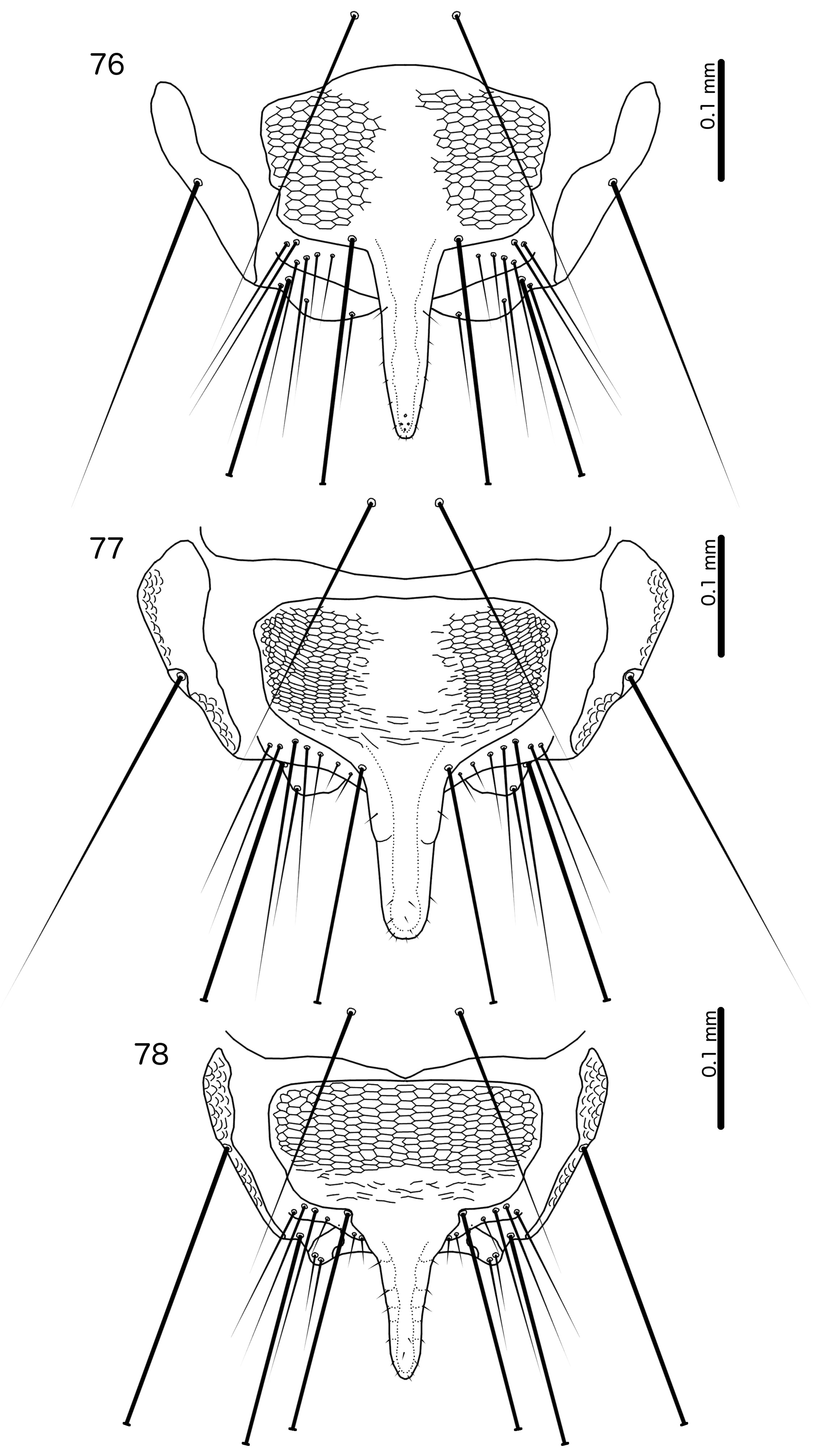
( Figs 1–2 View FIGURES 1–2 , 21 View FIGURES 21–24 , 33–35 View FIGURES 33–44 , 69 View FIGURES 69–70 , 76 View FIGURES 76–78 , 88 View FIGURE 88 )
Oxylipeurus himalayensis burmeisteri ( Taschenberg, 1882) ; Clay 1938: 169 (in partim).
Reticulipeurus himalayensis burmeisteri (Tasch.) ; Złotorzycka 1966: 112 (in partim).
Oxylipeurus burmeisteri ( Taschenberg, 1882) ; Price et al. 2003: 202 (in partim).
Type host. Lophophorus sclateri sclateri Jerdon, 1870 —Sclater’s monal.
Type locality. Yunnan Province, China .
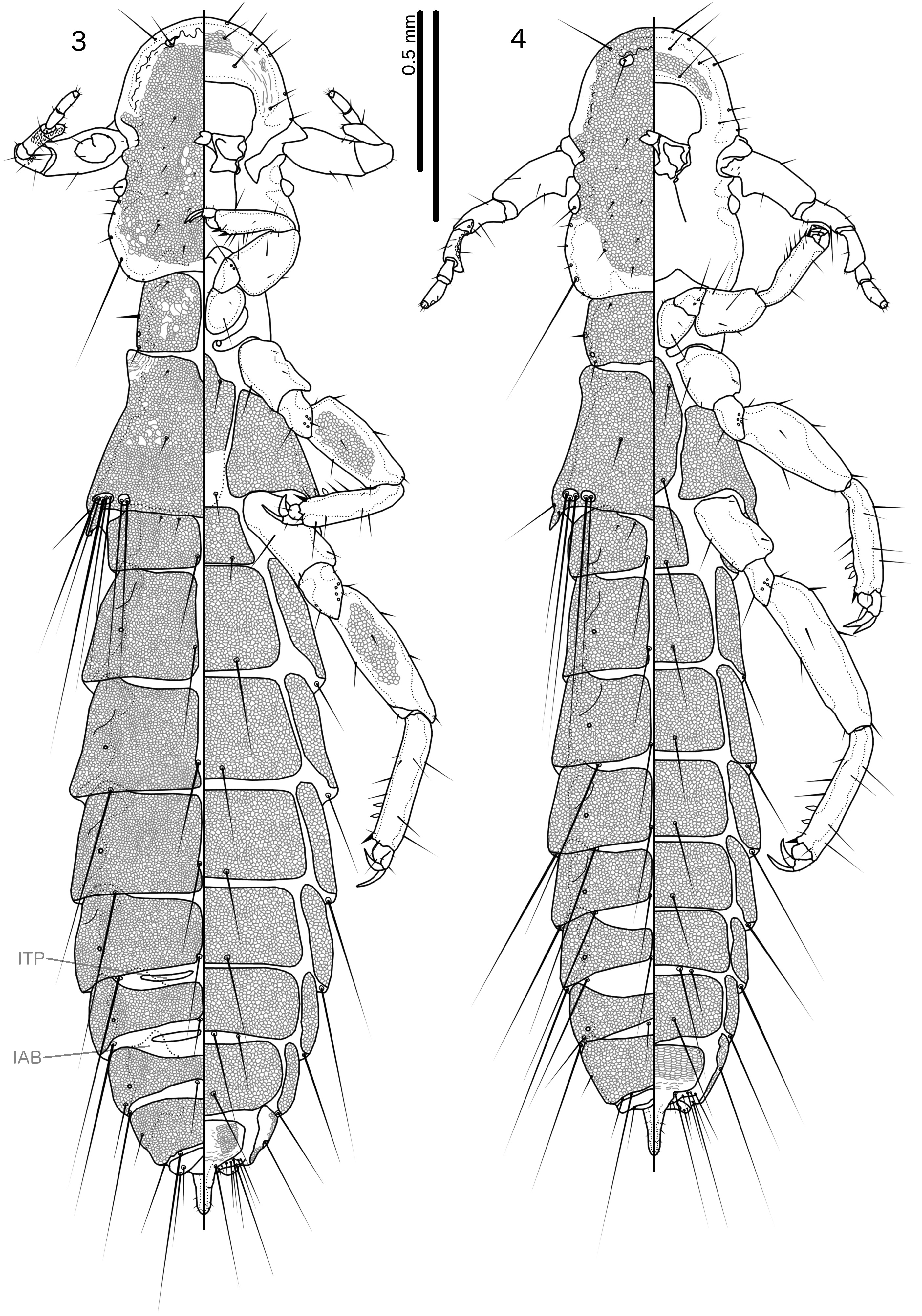
Diagnosis. Cataphractomimus impervius is most similar to C. mirapelta , based on the larger size of these two species compared to the other species in the genus ( Table 1 View TABLE 1 ; C. burmeisteri is also smaller), as well as their proportionately larger scapes (compare Figs 21–22 View FIGURES 21–24 with Fig. 23 View FIGURES 21–24 ). However, C. impervius and C. mirapelta can be separated by the following characters: Inter-tergal plates absent in male C. impervius ( Fig. 1 View FIGURES 1–2 ) but present in male C. mirapelta ( Fig. 3 View FIGURES 3–4 ); male tergopleurite XI medianly separate in C. impervius ( Fig. 1 View FIGURES 1–2 ) but medianly continuous in C. mirapelta ( Fig. 3 View FIGURES 3–4 ); subgenital plate proportionately narrower and with a more slender and gently tapering stylus in C. impervius ( Fig. 76 View FIGURES 76–78 ) than in C. mirapelta ( Fig. 77 View FIGURES 76–78 ); rugose area of male antennae more extensive in C. mirapelta ( Fig. 22 View FIGURES 21–24 ) than in C. impervius ( Fig. 21 View FIGURES 21–24 ); ventral sclerite of male mesosome clearly delimited laterally and with roughly triangular shape in C. impervius ( Fig. 35 View FIGURES 33–44 ) but diffuse laterally and with more or less flat anterior margin in C. mirapelta ( Fig. 38 View FIGURES 33–44 ); parameres longer and more slender in C. impervius ( Fig. 34 View FIGURES 33–44 ) than in C. mirapelta ( Fig. 37 View FIGURES 33–44 ); antero-lateral extensions of mesosome with smoothly rounded anterior margins in C. impervius ( Fig. 35 View FIGURES 33–44 ), but with more angular anterior margin in C. mirapelta ( Fig. 38 View FIGURES 33–44 ).
Description.
Both sexes. Head shape, structure, reticulation, and chaetotaxy as in Figs 1–2 View FIGURES 1–2 ; dorsal preantennal suture not visible in examined specimens. Marginal carina narrow. Head sensillus s5 situated about as far from pns as from s3. Thoracic and abdominal plates, chaetotaxy and approximate reticulation patterns as in Figs 1–2 View FIGURES 1–2 . Pronotum with deep anterior incision along midline ( Fig. 1 View FIGURES 1–2 ). Inter-tergal plates and internal anterior bulges absent. Tergopleurite XI medianly separate.
Male. Antenna as in Fig. 21 View FIGURES 21–24 ; rugose area limited to flagellomere I. Subgenital plate formed from sternal plate IX+X only, separate from sternal plate VIII ( Fig. 76 View FIGURES 76–78 ); reticulation of subgenital plate incomplete medianly; stylus slender ( Fig. 76 View FIGURES 76–78 ). No lateral sternal plates on abdominal segment XI. Basal apodeme as in Fig. 33 View FIGURES 33–44 , widening gradually towards distal end. Ventral sclerite of mesosome roughly triangular, separate from mesosome, with several small pores visible near lateral ends ( Fig. 35 View FIGURES 33–44 ). Rugose area of mesosome extensive. Antero-lateral extensions of mesosome with smooth anterior margins. Parameres elongated, slender ( Fig. 34 View FIGURES 33–44 ). Measurements as in Table 1 View TABLE 1 .
Female. Antennae as in Fig. 2 View FIGURES 1–2 . Subgenital plate medianly continuous ( Fig. 69 View FIGURES 69–70 ), but with deep anterior incision along midline. Lateral accessory vulval plates with 1–2 microsetae proximally on each side; vulval margin with 13– 17 marginal and submarginal setae on each side (exact position often hard to establish, and therefore here counted together). Subvulval plates slender, not extending much below vulval margin. Measurements as in Table 1 View TABLE 1 .
Etymology. The specific epithet is from “ impervius ”, Latin for “impassable”, referring to the extensive “armour” of the species.
Type material. Holotype ♂, Yunnan [ China], May 1906, R. Meinertzhagen , 3626, NHMUK010682345 View Materials ( NHML) [only specimen on slide with intact stylus and antennae, lower row, marked with red dot on slide] . Paratypes: 6♂, 8♀, same data as holotype, NHMUK010682343 View Materials , NHMUK010682345–6 View Materials ( NHML) .



| R |
Departamento de Geologia, Universidad de Chile |
| NHML |
Natural History Museum, Tripoli |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cataphractomimus impervius
| Gustafsson, Daniel R., Lei, Lujia, Chu, Xingzhi & Zou, Fasheng 2020 |
Oxylipeurus burmeisteri ( Taschenberg, 1882 )
| Price, R. D. & Hellenthal, R. A. & Palma, R. L. & Johnson, K. P. & Clayton, D. H. 2003: 202 |
Reticulipeurus himalayensis burmeisteri (Tasch.)
| Zlotorzycka, J. 1966: 112 |
Oxylipeurus himalayensis burmeisteri ( Taschenberg, 1882 )
| Clay, T. 1938: 169 |