
Transeius herbarius ( Wainstein, 1960 )
|
publication ID |
https://doi.org/ 10.15407/zoo2023.03.191 |
|
publication LSID |
lsid:zoobank.org:pub:0DF130DB-45F9-423F-A95B-489147F5BA79 |
|
persistent identifier |
https://treatment.plazi.org/id/03BADE00-1F26-FFE4-79CE-1F1BFB0D881C |
|
treatment provided by |
Felipe |
|
scientific name |
Transeius herbarius ( Wainstein, 1960 ) |
| status |
|
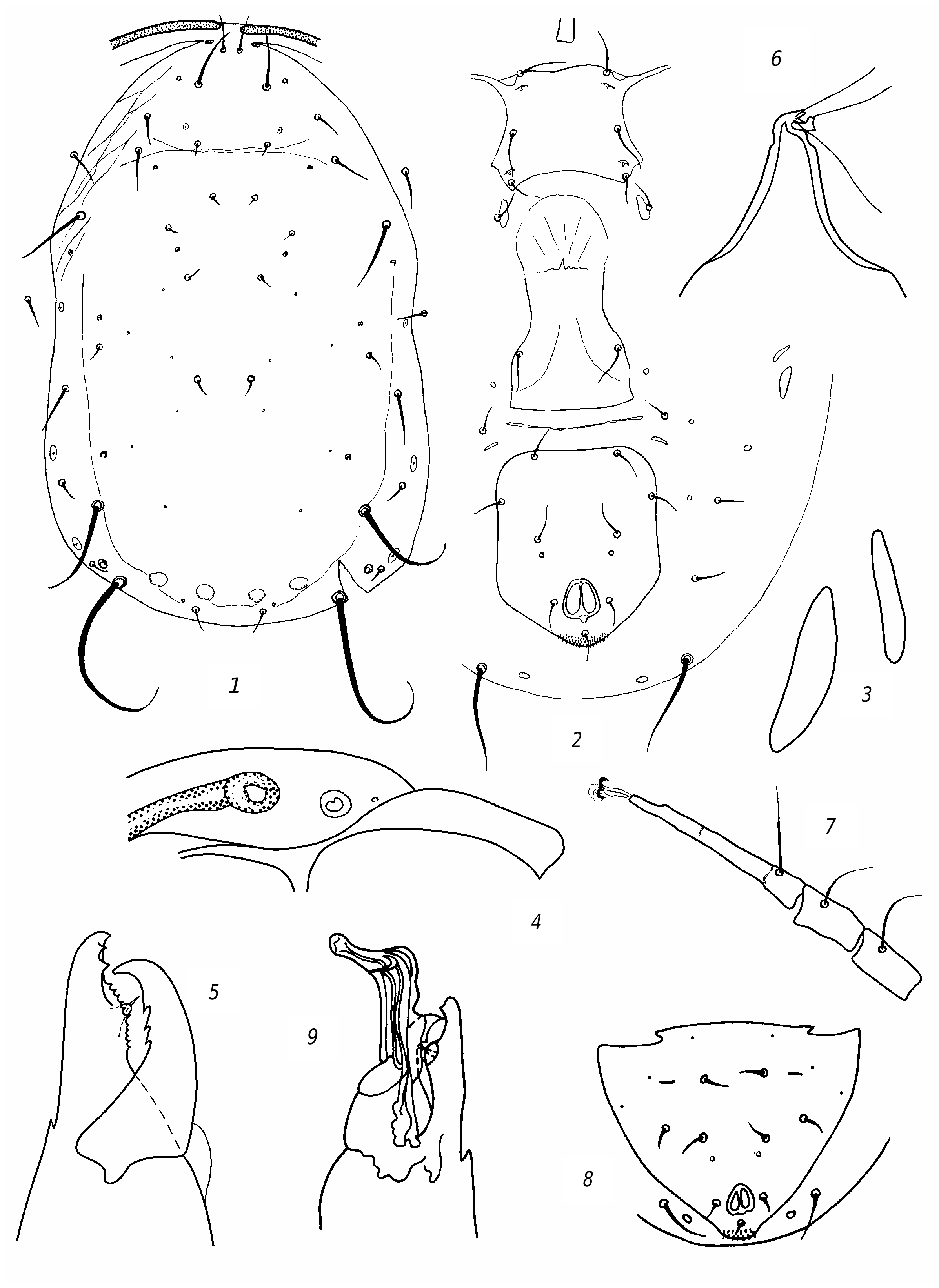
Transeius herbarius ( Wainstein, 1960) View in CoL ( fig. 3 View Fig )
Typhlodromus herbarius Wainstein, 1960: 687 ; Hirschmann, 1962: 225;
Amblyseius (Amblyseius) herbarius: Ehara, 1966: 14 ;
Amblyseius herbarius: Kolodochka, 1973: 80 ; 1974 a: 88; 1978: 40; Arutunjan, 1977: 35; Wainstein, 1977: 236; Begljarov, 1981: 33, 74; Kolodochka & Omeri, 2011: 44;
Neoseiulus herbarius: Moraes et al., 1986: 82 ;
Transeius herbarius: Chant & McMurtry, 2007: 71 View in CoL ; Faraji et al., 2011: 223.
Material. Type. Syntypes: 1 ♀, Kazakhstan, Astana , state farm, [leaf] litter, <under> raspberry, 18.08.1955, specimen #1015 ; 3 ♀, Almaty, Kargalinka, base of Plant Protection Station , on Agrimonia sp. , 19.04.1956, specimen #1368 ; 2 ♀, ibid., Carduus sp. , 04.05.1956, specimen #1422 ; 3 ♀, ibid., same data, specimen #1421 ( SIZK; B.A. Wainstein's collection) .
Non-type. 63 ♀, 9 Ơ (72 specimens): Ukraine, AR Crimea, Donetsk, Cherkasy, Chernihiv, Kherson, Kyiv, Luhansk, Rivne, Vinnytsia, Zhytomyr Region ( SIZK) .
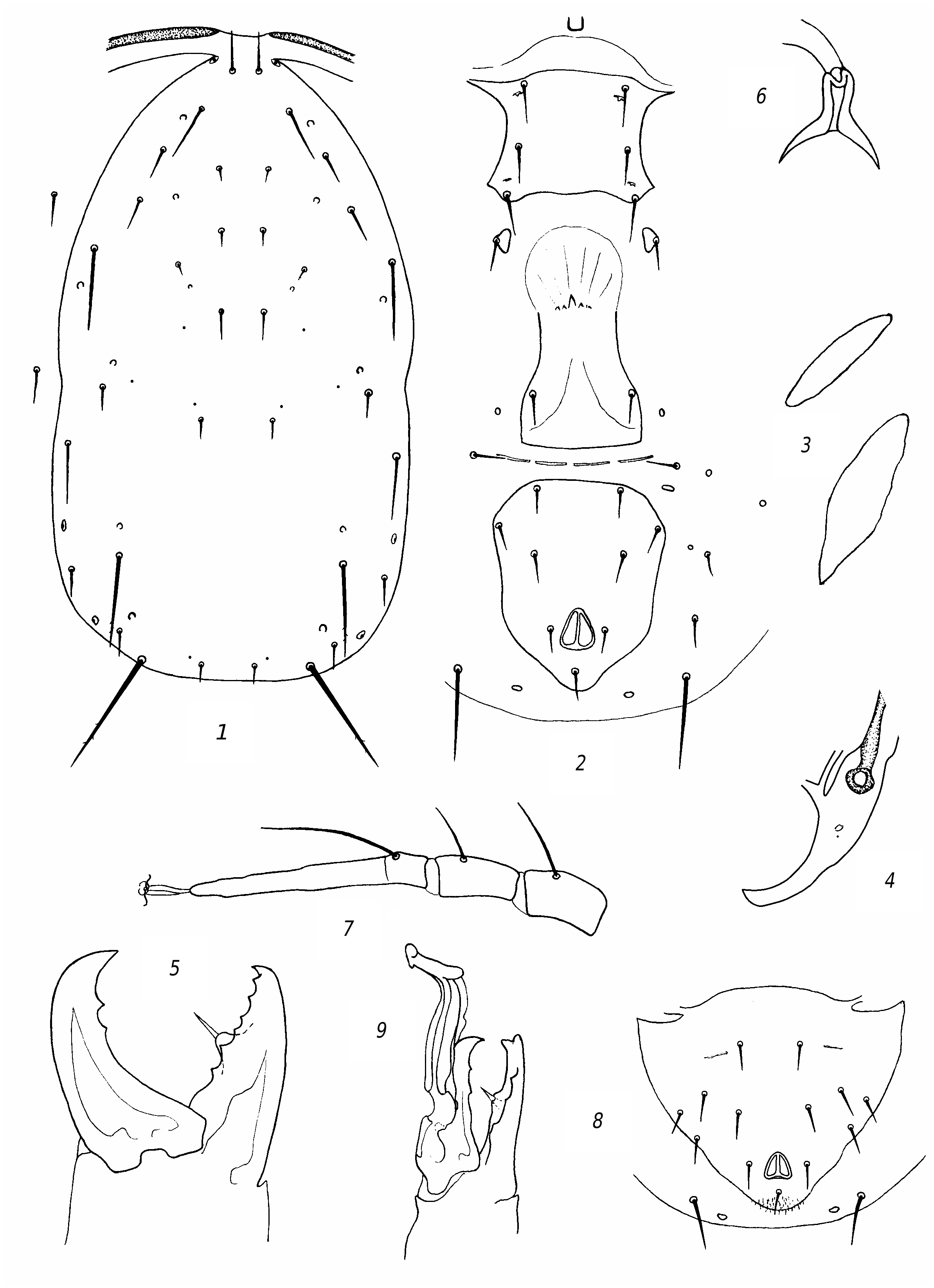
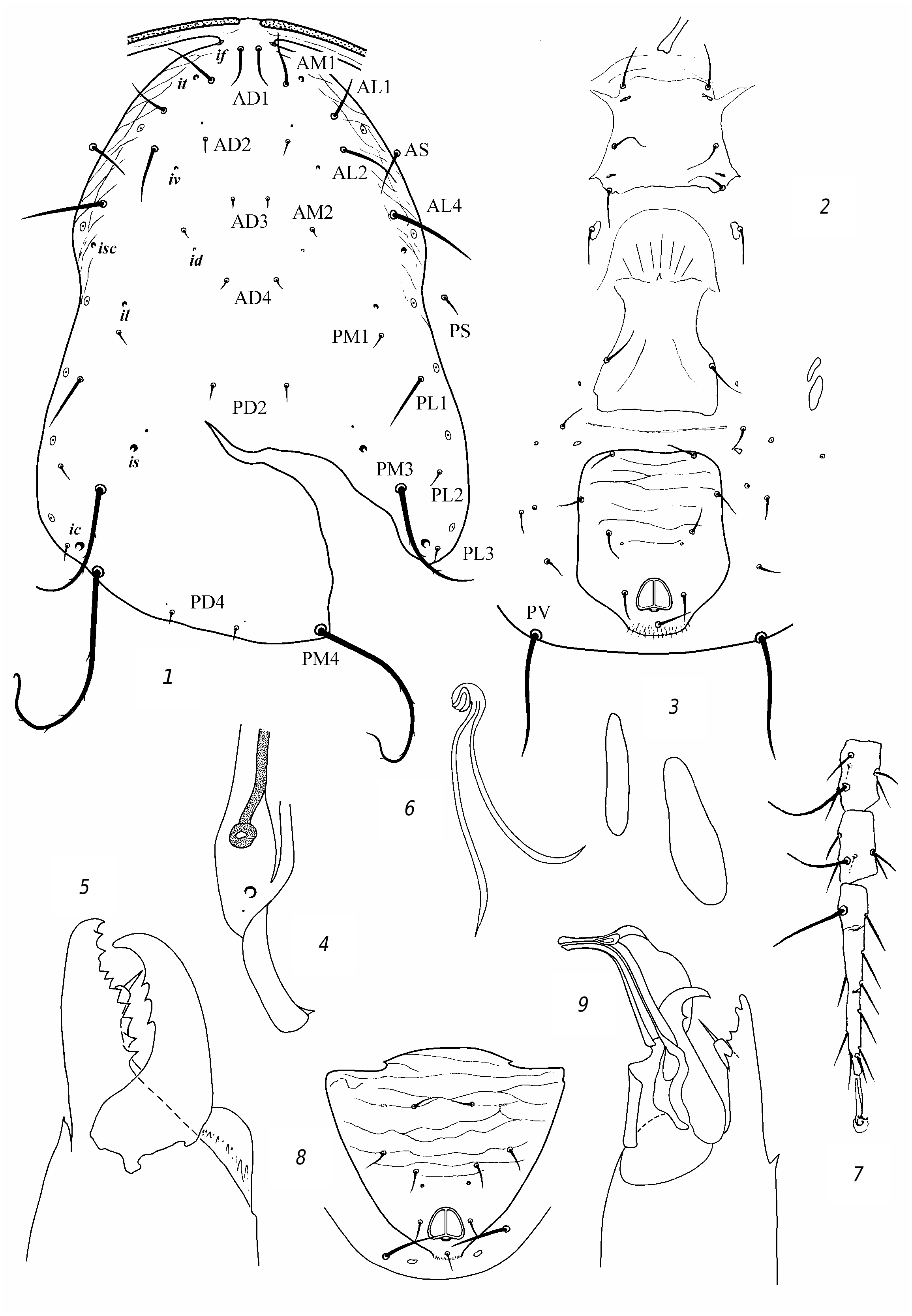
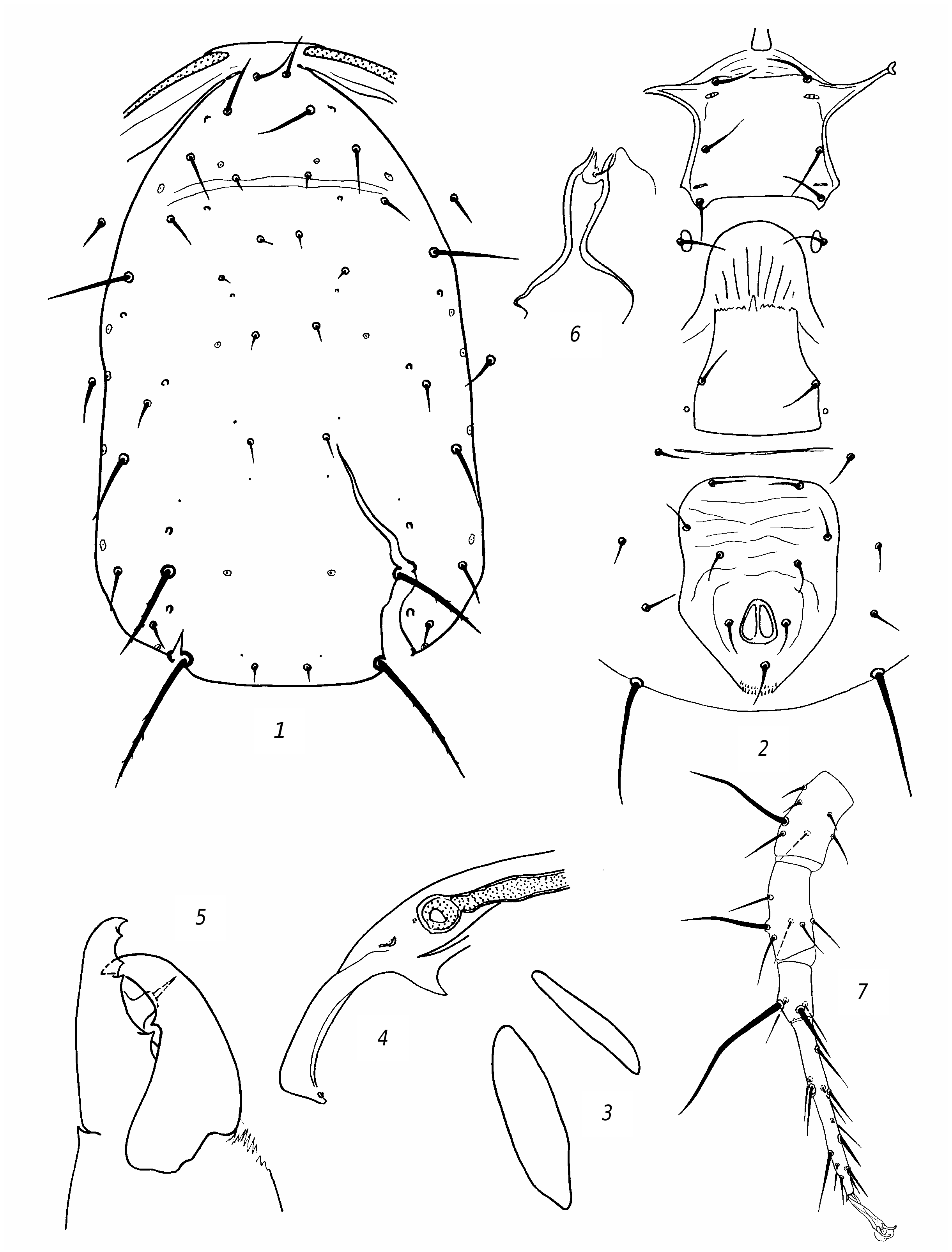
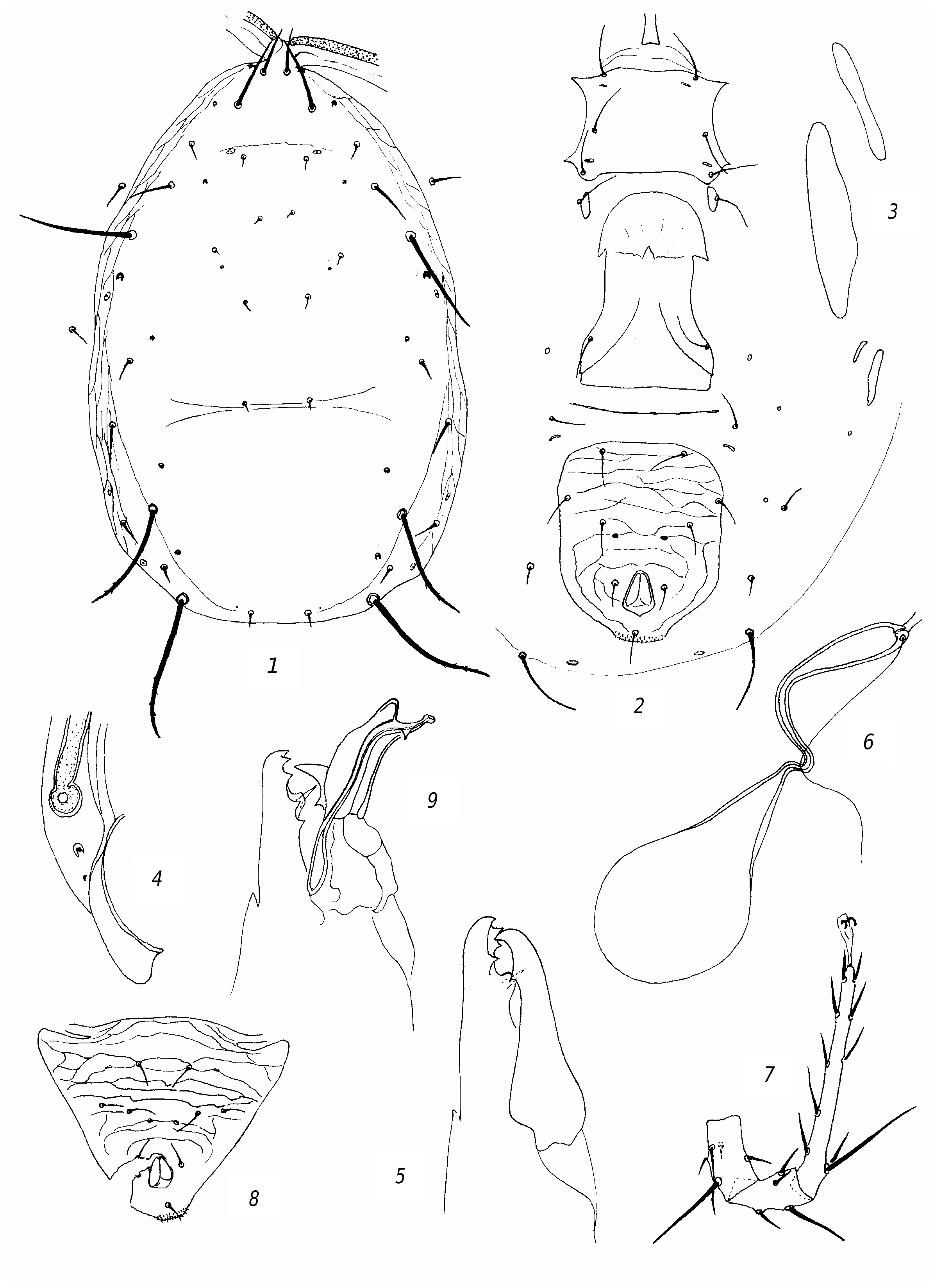
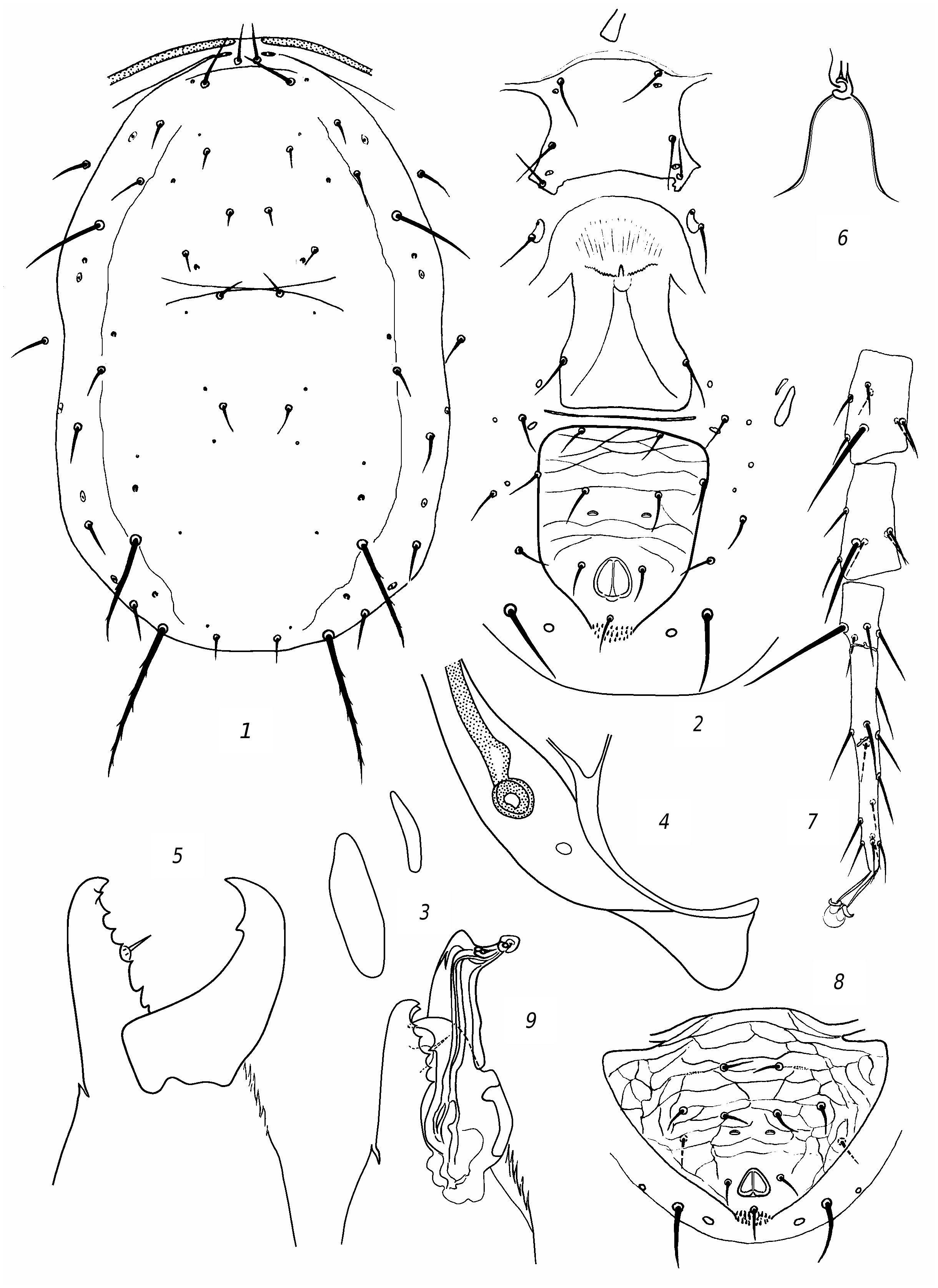
R e d e s c r i p t i o n. F e m a l e. Dorsal shield ( fig. 3 View Fig , 1 View Fig ) well sclerotized, mostly smooth, sometimes barely perceptibly reticulate, with slight lateral emarginations. Dorsal setae sharp, vary in length; solenostomes 7 pairs (it, iv, id, isc, il, is, ic). Setae PM3 and PM4 slightly serrated, rest smooth. Seta AM1 as long as distance to the theca of seta AL1 or slightly longer. Seta AL1 somewhat (3 µm) longer than AL3. Seta PL1 more than twice as long as setae PL2 and PL3. The peritreme do not extend beyond the level of the theca of seta AD1. Ventrianal shield with slight lateral emarginations, transversely striated; anal pores missing ( fig. 3 View Fig , 2 View Fig ). The posterior metapodal shield is fusiform, the anterior one is narrower and shorter nearly 1.5 times ( fig. 3, 3 View Fig ). Posterior part of peritremal shield curved, pointed ( fig. 3 View Fig , 4 View Fig ). Chelicera on Df with 6 teeth and on Dm with 2 ( fig. 3 View Fig , 5 View Fig ). The spermatheca is small, with thick wall of the funnel, thinning towards the sacculus; the massive atrium immersed into base of the funnel ( fig. 3 View Fig , 6 View Fig ). Leg IV with 3 macrosetae: the longest on the tarsus ( fig. 3 View Fig , 7 View Fig ). Genu and tibia III, genu and tibia II, as well genu I with one small macroseta.
Measurements: Lds 350, Wds 190, Lvas 128, Wvas 97, Ltar IV 131; setae length: AD1 25; AD2 7; AD3 8; AD4 11; PD2 11; PD4 8; AM1 33; АМ2 7; AL1 25; AL3 22; AL4 45; PL1 36; PL2 14; PL3 14; PM1 14; PM3 50; PM4 70; AS 22; PS 19; PV 47; MCh IV: ge 47, ti 36, ta 78; MCh III: ge 23, ti 23, MCh II: ge 23, ti 23; MCh I: ge 30.
M a l e. Preanal setae 5 pairs; anal pores missing ( fig. 3 View Fig , 6 View Fig ). The spermatodactyl is curved ( fig. 3 View Fig , 7 View Fig ). Lds 305.
Diagnosis. Transeius herbarius is very similar to T. fragilis (Kolodochka & Bondarenko) reliably differing from T. fragilis by short and thick wall funnel of spermatheca, more than 6 teeth on Df and on 2 on Dm (in T. fragilis , only 4 teeth on Df and 1 on Dm; the basal tooth on Df missing), and dorsal setae PL2, PM4, and PV shorter (14, 70, and 47 in T. herbarius , and 22, 85, and 65 in T. fragilis , respectively).
D i s t r i b u t i o n, h a b i t a t, o c c u r r e n c e. Europe, the Caucasus, the Middle East. Ukraine: steppe zone, forests-teppe zone, forest zone (Polissia), Crimea (second ridge of mountains), confined to herbaceous plants; common species.
N o t e s 1. Four of the 16 T. herbarius syntypes accessions that Dr. Wainstein dealt with were studied (listed above). Specimens #1422 (2 ♀), and #1423 (3 ♀) dried up and could not be studied. The rest of the specimens are missing from the collection.
2. Description, dimensions and illustrations are based on better preserved non-type specimens from Kyiv Region.
3. As has been previously established, T. herbarius mites reproduce under laboratory conditions in the manner of thelytoky (Kolodochka, 1974), i. e. only females develop from unfertilised eggs. Males are either unknown (obligate parthenogenesis) or appear under certain conditions (facultative parthenogenesis).
Subsequently, the study of earlier collections showed that T. herbarius may be reproduce in a bisexual manner under natural conditions, which is common to most phytoseiids ( Kolodochka, 1984 a, b). Because of the rarity of male specimens, the localities where they were collected are listed here: Kyiv, wasteland near the biological laboratory of the Kyiv State University [50°23'05.0" N 30°28'33.0" E], Arctium sp. , 08.07 1972, specimen #994, 2 ♀, 1 Ơ; ibid., same host under a solitary apple tree, 18.08 1970, specimen #1170, 8 ♀, 3 Ơ.
Later, males of N. herbarius were discovered as well in the material of field collections during two expeditions: in Ukraine: Crimea, Tarkhankut Peninsula, 7 km W from the vil. Chernomorskoe, Echium sp. , relic steppe territory, 23.05.1979, specimen #3359 b, 1 Ơ. Beyond Ukraine — Moldova: Calarash, private garden, Arctium sp. , 10.07.1978, specimen #2992, 1 Ơ; ibid., Cichorium sp. , 10.07.1978, specimen #2994a, 6 ♀ 1 Ơ, specimen #2994b, 7 ♀, 1 Ơ (Kolodochka).
For unclear reasons, this line of research has not yet attracted much attention from scientists involved in the search for effective predators for plant protection and has not been further developed. At the same time, it can be very useful in solving not only purely theoretical problems, but also have practical application in the development of new directions in the development of biological methods to protect plants from pests. An important fact is that the population growth of parthenogenetic predator species occurs with the full realisation of the generative potential of females, without the need to spend a significant part of it on the reproduction of males. If we consider that males can make up about a third of the individuals in a population (our own data) and that the rate of pest destruction becomes a decisive factor in the choice of the appropriate method of plant protection, parthenogenetic species gain a significant advantage over species with a bisexual mode of reproduction.

| SIZK |
Schmaulhausen Institute of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Transeius herbarius ( Wainstein, 1960 )
| Kolodochka, L. A. 2023 |
Transeius herbarius: Chant & McMurtry, 2007: 71
| Faraji, F. & Cobanoglu, S. & Cakmak, I. 2011: 223 |
| Chant, D. A. & McMurtry, J. A. 2007: 71 |
Neoseiulus herbarius:
| Moraes, G. J. & McMurtry, J. A. & Denmark, H. A. 1986: 82 |
Amblyseius herbarius:
| Kolodochka, L. A. & Omeri, I. D. 2011: 44 |
| Begljarov, G. A. 1981: 33 |
| Kolodochka, L. A. 1978: 40 |
| Arutunjan, E. S. 1977: 35 |
| Wainstein, B. A. 1977: 236 |
| Kolodochka, L. A. 1974: 88 |
| Kolodochka, L. A. 1973: 80 |
Amblyseius (Amblyseius) herbarius:
| Ehara, S. 1966: 14 |
Typhlodromus herbarius
| Hirschmann, W. 1962: 225 |
| Wainstein, B. A. 1960: 687 |