
Melanosuchus latrubessei, Souza-Filho & Guilherme & Toledo & Carvalho & Negri & Maciente & Cidade & Lacerda & Souza, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4894.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:19BD5D89-5C9C-4111-A271-98E819A03D8E |
|
DOI |
https://doi.org/10.5281/zenodo.4324167 |
|
persistent identifier |
https://treatment.plazi.org/id/03BB3D0B-FF86-FFFC-FF04-FE833390F86C |
|
treatment provided by |
Plazi |
|
scientific name |
Melanosuchus latrubessei |
| status |
sp. nov. |
Melanosuchus latrubessei sp. nov.
( Figure 2 View FIGURE 2 , 3 View FIGURE 3 , 4 View FIGURE 4 , 5 View FIGURE 5 and 6 View FIGURE 6 )
Zoobank species: urn:lsid:zoobank.org:act:BB6E5F62-348B-4880-BB42-423BA34E0EE4
Holotype. UFAC 2793 , right part of a rostrum.
Etymology. The specific epithet latrubessei is in honor to the Argentinean geomorphologist Dr. Edgardo M. Latrubesse. His geopaleontological contributions were very important to the increase of knowledge about origin and evolution of the Solimıes Formation. Dr. Edgardo also provided important scientific contributions for Universidade Federal do Acre through collaborations in partnership with Laboratório de Pesquisas Paleontológicas.
Diagnosis. Melanosuchus latrubessei possess the following autapomorphies: (1) the contact between maxillamaxilla is elongated and extends from the space between the second and third alveoli until to the space between the sixth and seventh alveoli; and (2) the rostral ridge 2 is well-developed and well-marked, being anteroposteriorly inclined, and composed exclusively by the maxillary bone.
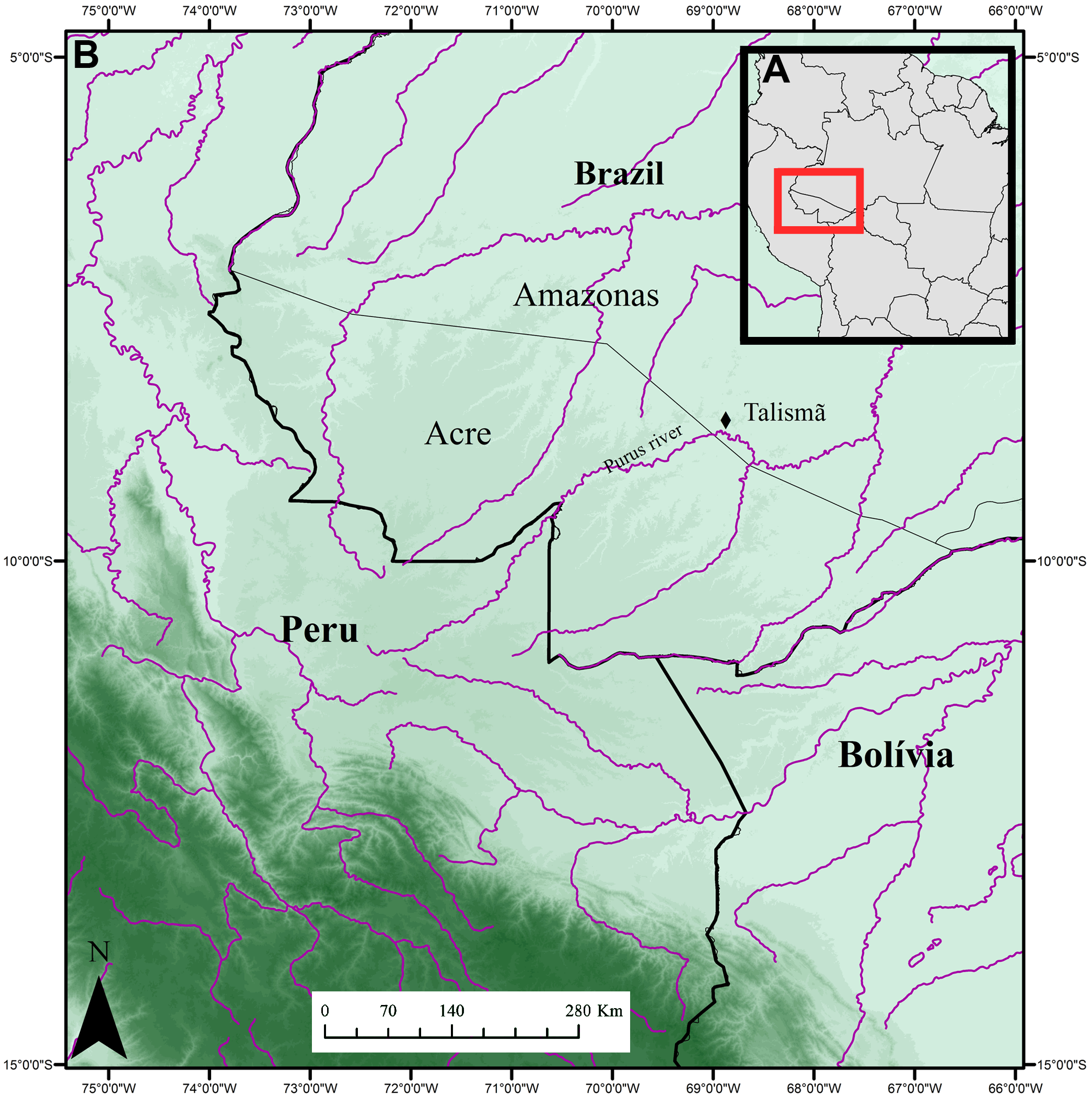
Occurrence. The Talism „ outcrop is located on the bank of the Purus River , in Southern region of the Amazonas State, Brazil (08°46’37,8”S; 68°54’15,1”W; Figure 1 View FIGURE 1 ). The outcrop has approximately eight meters of vertical length and the rocks are formed by fine grained silts and clays floodplain sediments, with secondary gypsum and carbonate veins. Presenting his maximum deposition 10.89 ± 0.13 Ma, being at least Tortonian in age (Bissaro- Júnior et al. 2019) GoogleMaps .
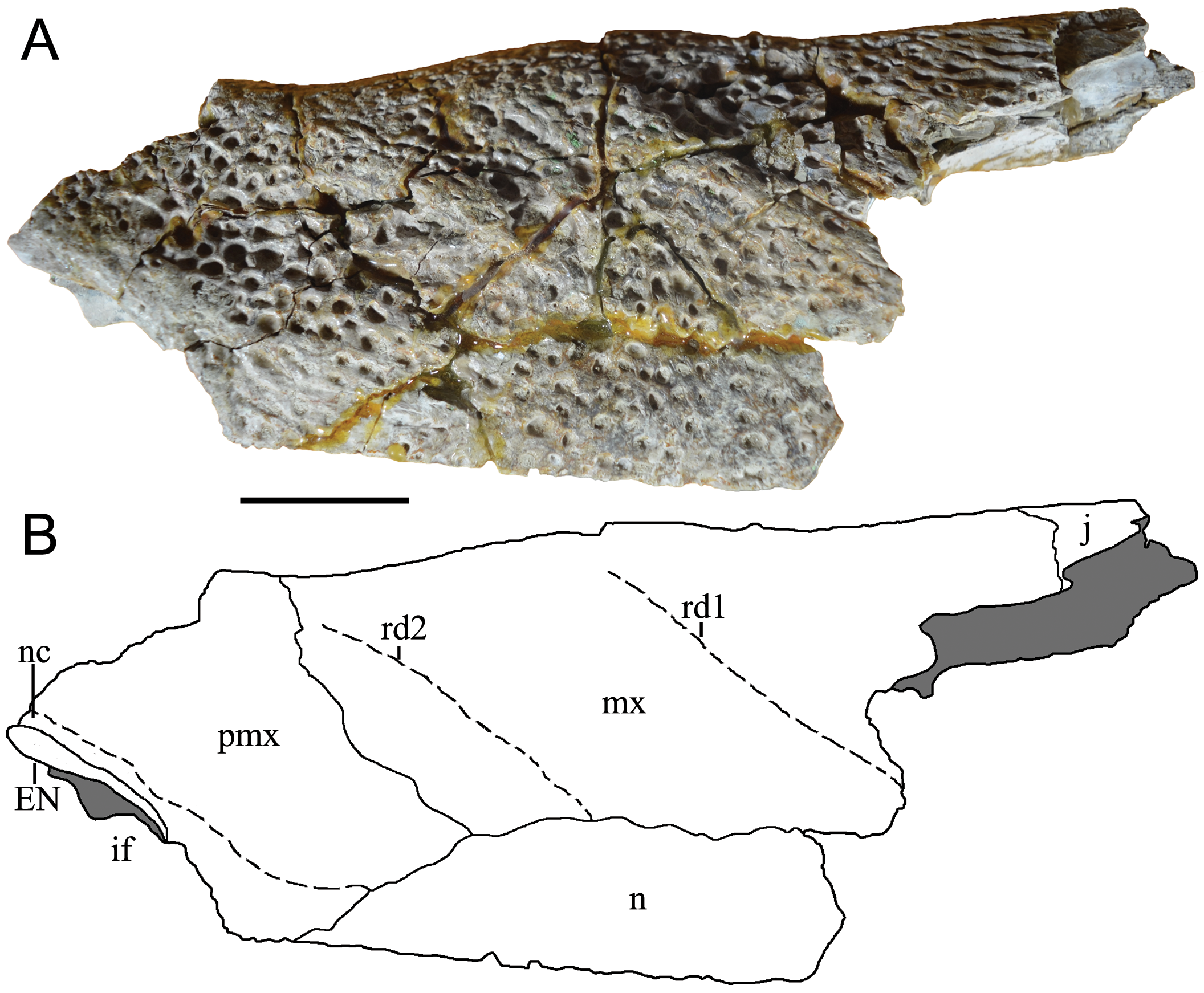
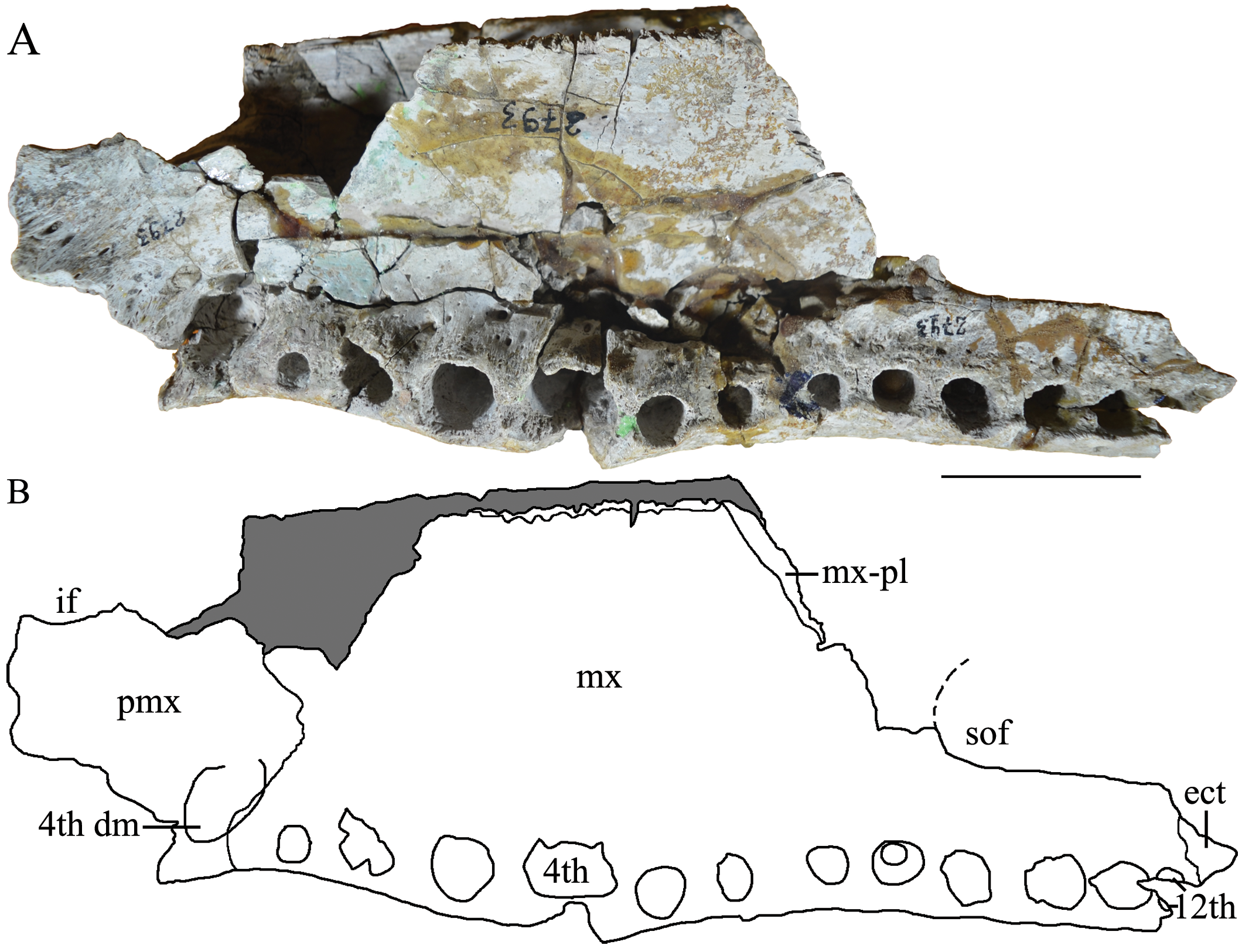
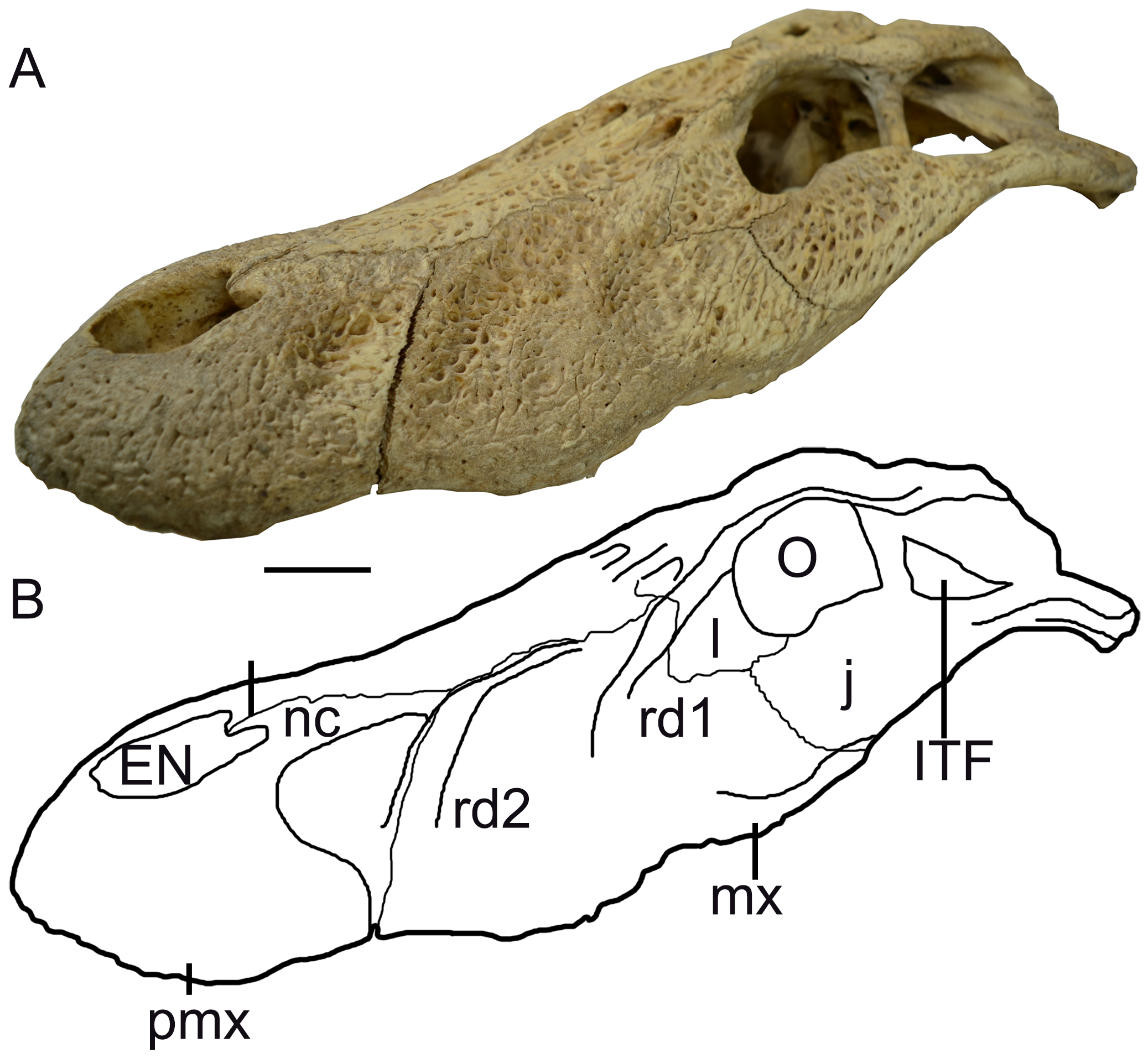
Description. The skull fragment has a maximum length of 287 mm and maximum width of 87.7 mm (measured at the level of third maxillary alveolus). The holotype of M. latrubessei, UFAC 2793 , consists on the right half of the rostrum, with only the posterior portion of the premaxilla preserved, almost complete maxilla and the nasal bone lacking the most posterior region ( Figure 2 View FIGURE 2 , 3 View FIGURE 3 and 4 View FIGURE 4 ). A small fragment of the right ectopterygoid is also preserved and articulated with the posterior portion of the maxilla ( Figure 3 View FIGURE 3 ). The premaxilla lacks most of the lateral and anterior region; therefore, no alveoli is preserved. In dorsal view, the premaxilla, preserves the right lateral and posterior portions of the external nares ( Figure 2 View FIGURE 2 ) and, in ventral view, preserves only the right lateral margin of incisive foramen ( Figure 3 View FIGURE 3 ). The maxilla has preserved the joint surface with the jugal, which can be seen in the dorsal and lateral views ( Figure 2 View FIGURE 2 and 4 View FIGURE 4 ). In ventral view, the maxilla presents, in its posterior middle corner, the articular surface of the anterior process of the palatine and on the lateral margin, twelve alveoli ( Figure 4 View FIGURE 4 ). No diagenetic torsions or modifications, beside those of the broken portions, are observed.
M. latrubessei UFAC 2793 presents a well-marked bone ornamentation uniformly distributed along the entire dorsal surface of the rostrum, with exception of the region around the external nares ( Figure 2 View FIGURE 2 and 4 View FIGURE 4 ). The ornamentation consists of irregular but usually round and well-marked pits, with small variations in size. The external nares have only the posterior margin and the posterior lateral margin preserved ( Figure 2 View FIGURE 2 ). The posterior margin, although not complete, appears to form an almost straight line, with a shallow convexity at the transition to the lateral margins. The lateral margin opens laterally, with the anterior portion more lateralized than the posterior, this inclination makes the anterior margin of the external nares wider than the posterior one ( Figure 2 View FIGURE 2 ).
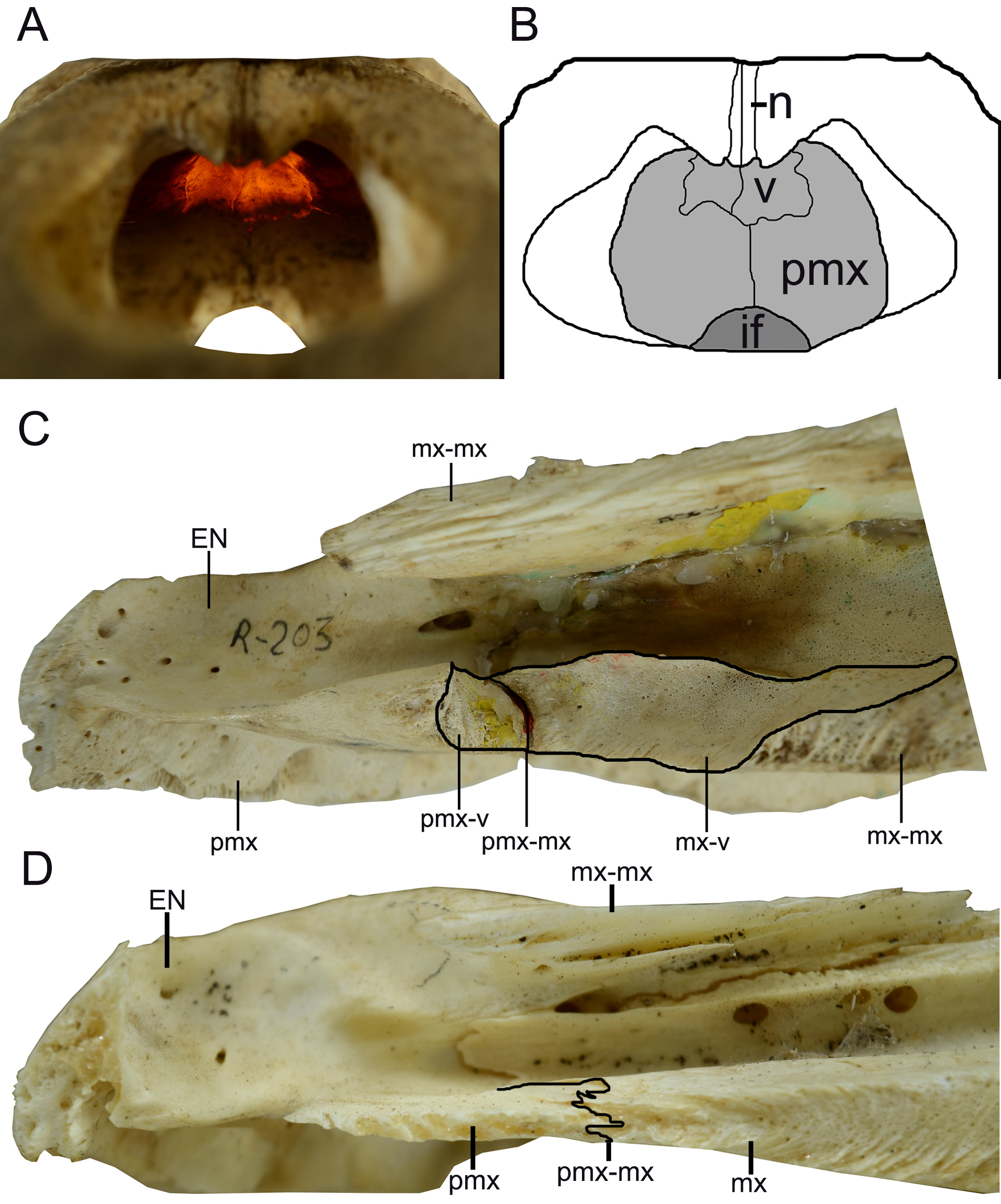
Only the right lateral middle portion of the incisive foramen is preserved, this portion represents a shallow concavity that is in the posterior portion of the premaxilla ( Figure 3 View FIGURE 3 ). However, in the absence of premaxillary alveoli and the fragmentary condition of the structure, the exact location and shape of the incisive foramen can only be tentatively inferred. This shallow concavity seems to be related to an anteroposteriorly long incisive foramen, which probably extends from the last and penultimate premaxillary alveoli to, at least, the third premaxillary alveoli (considering five premaxillary alveoli, found in Crocodylia—Brochu 1999, 2011, 2013). However, the shape of incisive foramen as a whole cannot be inferred.
The right suborbital fenestra is partially preserved, and only the portions that contribute to the maxilla are present, with the anterior portion of the lateral margin and the lateral portion of the anterior margin ( Figure 3 View FIGURE 3 ). The preserved anterior margin is a shallow concavity and the lateral margin is straight and slightly inclined mediolaterally, with the anterior portion more medial than the posterior one ( Figure 3 View FIGURE 3 ). The maxillary margin of the suborbital fenestra is straight, without any projection in the fenestra; moreover, the anterior margin of suborbital fenestra reaches the space between the ninth and the eighth alveoli ( Figure 3 View FIGURE 3 ).
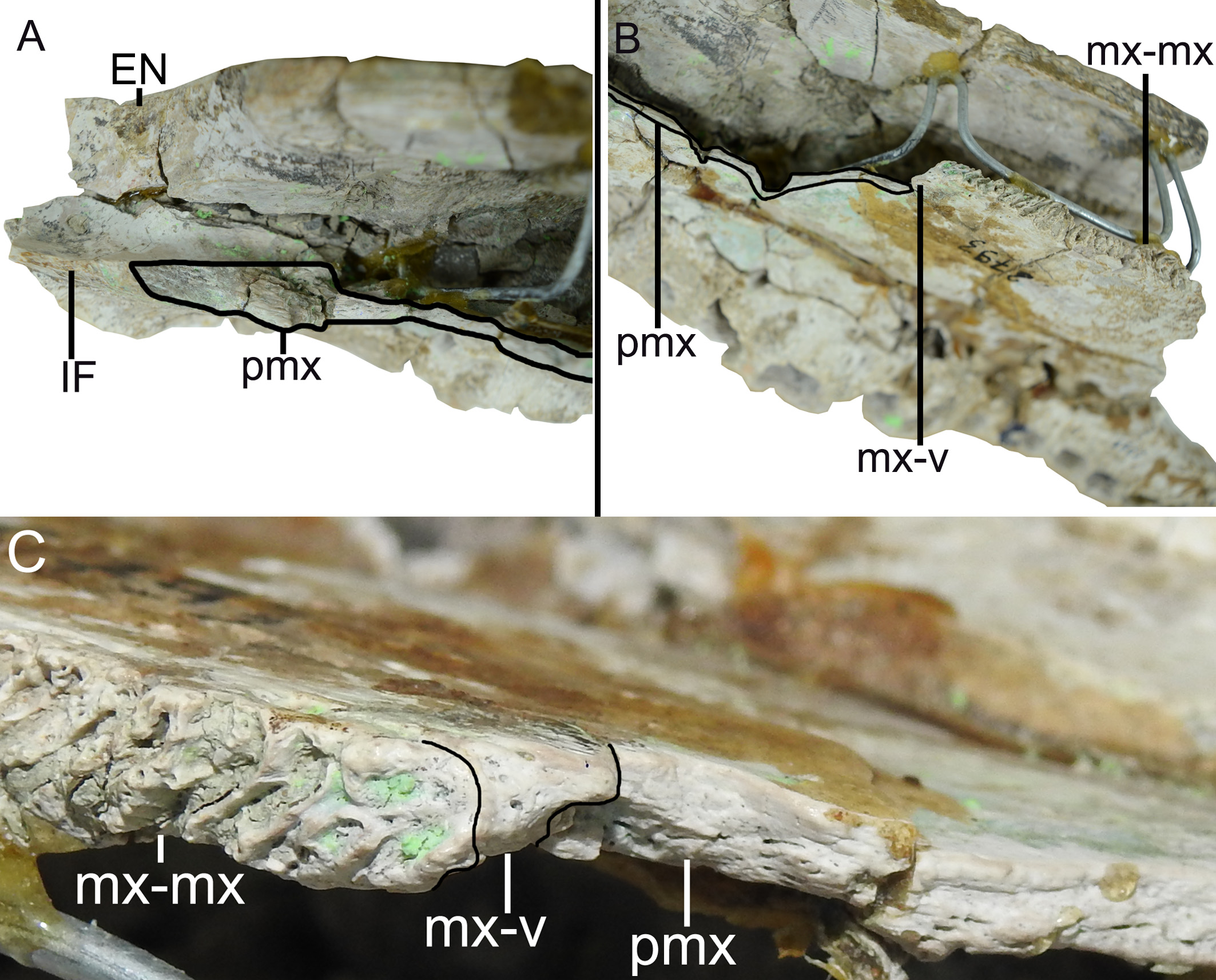
The premaxilla, in dorsal view, has a posterior wedge-shaped process that extends until the position of the third maxillary alveolus. This posterior process has the apex aligned with the suture between the maxillary-nasal, being the lateral portion of the premaxilla posterior process sutured with the maxilla, in the most anterior part, and medially with the nasal bone ( Figure 2 View FIGURE 2 and 4 View FIGURE 4 ). The premaxilla has a continuous crest delimiting the lateral and posterior limits of the external nares, which is here referred as narial crest ( Figure 2 View FIGURE 2 and 4 View FIGURE 4 ). It is important to note that, in dorsal view, the right premaxilla sutures with the left premaxilla, at least dorsally, with this contact being located immediately posterior to the margin of the external nares ( Figure 2 View FIGURE 2 ). The premaxilla-premaxilla contact in dorsal view can be better evidenced in a sagittal cut, where the medial surface of the right premaxilla and right nasal can be observed ( Figure 5 View FIGURE 5 ). The posterior margin of the external nares, composed entirely by the premaxilla, is preserved and does not have sutures with the nasal bones ( Figure 5 a, b View FIGURE 5 ).
The anterior portion of the nasal bone is broken, immediately above the anterior portion of the premaxilla. However the premaxilla is complete, indicating an overlapping suture (i.e. at least at this anterior portion the premaxilla lies over the nasal) ( Figure 5 a, d View FIGURE 5 ). This relation between premaxilla and nasal can be better understood due to the break of the nasal in ventral view, which evidences a contact between those bones and not a fusion demarcating a suture ( Figure 5 c View FIGURE 5 ). Moreover, the medial surface of the premaxilla is rugose as expected to a premaxilla-premaxilla contact, while the nasal bones present a horizontal and parallel lines, as expected in nasal-nasal contact ( Figure 5 d View FIGURE 5 ). The nasal bone is broad in the lateromedial direction, with its anterior wedge-shaped projection. The premaxillarymaxillary suture contacts the lateral limit of the nasal at the point where the wedge-shaped process begins ( Figure 2 View FIGURE 2 ).
The maxilla, in dorsal view, contacts anteriorly with the premaxilla and nasal, while having exclusive medial contact with the nasal bone ( Figure 2 View FIGURE 2 and 4 View FIGURE 4 ). There are a total of twelve maxillary alveoli. The longitudinal measurements of each alveolus, from the anterior most to the posterior most are, respectively (in mm): 6.88; 11.33; 14.75; 17.95; 9.58; 7.65; 7.29; 9.46; 11.32; 11.36; 10.9; and 8.19. The eighth alveolus has a preserved tooth, which did not have a complete dental eruption in vivo; therefore, the non-exposure of this tooth prevents a detailed description of its morphology. However, the apex of the tooth is visible and represents a convex structure.
The lateral border of maxilla, in dorsal and ventral views, has the anterior (position anteriorly to the first maxillary alveolus) and posterior (position of the eleventh and twelfth maxillary alveoli) portions projecting laterally. At the level of the fourth alveolus, in both dorsal and ventral views, there is a well-developed convexity ( Figure 2 View FIGURE 2 and 4 View FIGURE 4 ). In lateral view, the ventral margin of the maxilla has a well-marked concavity anteriorly to the convexity at the level of fourth maxillary alveolus, and posterior to this convexity there is another shallow concavity ( Figure 4 View FIGURE 4 ). In the posterior region of the maxilla, there is a surface corresponding to the contact between maxilla-jugal, where the jugal bone overlies the maxilla ( Figure 2 View FIGURE 2 and 4 View FIGURE 4 ). However, the jugal is not preserved.
The maxilla also has two well-developed rostral ridges ( Figure 2 View FIGURE 2 and 4 View FIGURE 4 ). The first (rostral ridge 1) is associated with the high development of the canthi rostralii; thus, although the interorbital region is not preserved, the presence of a well-developed canthi rostralii is proposed. Therefore, this difference in saturation surface reinforces the idea of a premaxilla-premaxilla contact exposed in dorsal view. However, as the nasal is not entirely preserved, we cannot rule out the possibility of nasal bones with a continuous contribution with the anterior process within the external nares, passing bellow the premaxilla-premaxilla contact ( Figure 5 d View FIGURE 5 ). In ventral view, the premaxilla-maxilla suture passes within the tooth occlusal pit. From the fourth dentary, and posteriorly to this excavation the suture has a small posterior wedge-shaped process that extend to the posterior end of the first maxillary alveoli ( Figure 3 View FIGURE 3 ). The other rostral ridge (rostral ridge 2) is entirely located within the maxilla next to the premaxilla-maxilla suture, in dorsal view ( Figure 2 View FIGURE 2 and 4 View FIGURE 4 ), and represents a dorsoventrally inclined bone intumescence, with the anterior portion ventral to the posterior ( Figure 4 View FIGURE 4 ). In ventral view, the maxilla-maxilla suture extends straight from the third to the sixth alveolus ( Figure 3 View FIGURE 3 ). Prior to the maxilla-maxilla suture, there is a smooth maxillary portion that opens slightly laterally and is here interpreted as the contact surface with the vomer. At the posterior limit of the maxilla, immediately behind the intermaxillary suture, there is the sutural region of the palatine, which is evidenced by a surface with thin crests. Thus, the palatine extends anteriorly to the level of the sixth alveoli ( Figure 3 View FIGURE 3 ).
At the posterior limit of the maxilla in ventral view, immediately posterior to the tooth row, there is the maxillaectopterygoid contact, with a small portion of the anterior region of the ectopterygoid being preserved ( Figure 3 View FIGURE 3 ). The ectopterygoid does not form the medial margin of any maxillary alveolus and the anterior process of this structure does not exceed the twelfth maxillary alveolus ( Figure 3 View FIGURE 3 ). The lateral portion of the maxilla to the suborbital fenestra is wide, being at least twice the width of the tenth alveolus ( Figure 3 View FIGURE 3 ).
Despite the fact that the vomer is not preserved and the region between the maxilla and premaxilla is severely damaged, there are some clear evidences in both the premaxilla and the maxilla of the presence of the vomer, which will be presented below. The first evidence is the presence of the aforementioned smooth anterior surface to the end of the maxilla-maxilla suture, which opens laterally and corresponds to the articular surface between the maxilla and the vomer ( Figure 2 View FIGURE 2 and Figure 6 b, c View FIGURE 6 ). The second evidence can be seen in medial view, due to the sagittal cut where the ventral portion of premaxilla has an intumescent region, which is broken but clearly detectable, with this structure being correspondent to the posterior portion of the premaxilla that contacts the anterior limit of the vomer ( Figure 6 a View FIGURE 6 ). Additionally, it is important to note that the broken pattern observed in the transition zone between premaxilla and maxilla can be expected for a specimen with an exposed vomer, as the presence of the vomer creates a fragilized zone in this portion of the rostrum.
Comparisons. Melanosuchus latrubessei shares with Melanosuchus niger the presence of a vomer that is exposed and located between the premaxilla-maxilla suture, as well as the well-developed rostral ridges, mainly the second and most anterior one. Other similarities worth mentioning here are: (1) the external nares with a keyholeshaped or circular format; (2) the premaxilla which has a right and left posterior processes, in ventral view, that does not surpass posteriorly the first maxillary alveoli; (3) the premaxilla-premaxilla contact at the dorsal surface immediately behind the posterior margin of the external nares, which represents an intraspecific variation within M. niger ; that can have the nasal bones reaching the external nares and being exposed dorsally inhibiting the premaxilla-premaxilla contact at this region; (4) the palatine, which projects well anterior to the end of the suborbital fenestrae and has an expanded anterior end; and, (5) the anterior margin of the suborbital fenestrae reaches the space between the eighth and ninth maxillary alveoli (differing from MCNC 243 specimen - former M. fisheri , which reaches the space between ninth and tenth alveoli).
The vomer exposure on the palate between premaxilla and maxilla is a characteristic of M. niger (e.g. Mook 1921; Medem 1963; Iordansky 1973; Brochu 1999; Bona et al. 2017b), and this feature was used as diagnosis to assign the fossil specimens to this genus (e.g. Medina 1976). However, the bone has never been properly described, even in detailed works on M. niger (e.g. Mook 1921; Medem 1963; Iordansky 1973; Medina 1976; Brochu 1999; Vieira et al. 2016; Bona et al. 2017b). In this work, we recognize the vomer as a hollow paired bone, located between the premaxilla and maxilla, in the medial and anterior region of the rostrum, with the paired bones being attached only by a contact, and not by a real suture. This can be evidenced by the thin projections from the maxillae, which overlap ventrally and dorsally the posterior and lateral limits of the vomer ( Figure 7 View FIGURE 7 ). The anterior portion of the vomer attaches, without suturing, to the posterior premaxillary intumescence ( Figure 7 View FIGURE 7 ).
The hollowness of the vomer, together with its weak contact towards the premaxilla and maxilla, make this bone and the surrounding area more fragile when the animal is dead; therefore, it is common the loss of the vomer with the break and/or disarticulation of associated bones, as observed in M. latrubessei ( Figure 3 View FIGURE 3 ; this is also evidenced in the studied M. niger , see Table 1). However, we reinforce that the existence of the vomer in M. latrubessei is supported by two features: (1) the anterior end of maxilla-maxilla suture at the palate culminates in a smooth maxillary surface that opens laterally ( Figure 6 b, c View FIGURE 6 ); and, (2) the premaxilla, in medial view from a sagittal cut, has at the posterior portion of the ventral surface an intumescence that was broken in the M. latrubessei specimen, but the characteristic brands remained ( Figure 6 a View FIGURE 6 ). The condition observed in M. latrubessei diverges from that seen in Caiman genus, which has the vomer restricted to the narial passage and located posteriorly, aligned with the prefrontals (e.g. Bona & Desojo 2011). It also diverges from Paleosuchus palpebrosus (Cuvier 1807) and the Caiman species by exhibiting premaxillae that are thicker and more robust than the maxillae ( Figure 7 c, d View FIGURE 7 ). For vomer condition in Alligator see Rieppel (1993).
Despite the aforementioned similarities shared between M. latrubessei and M. niger , they also present several differences that should be highlighted, such as: (1) M. latrubessei presents twelve maxillary alveoli, while M. niger has thirteen (including MCNC 243); (2) the narial crest in M. latrubessei is restricted to the border of the external nares and is isolated from the rostral ridges ( Figure 4 View FIGURE 4 ), while in M. niger the narial crest at the posterior corner of external nares has a posterior projection bordering the premaxilla-nasal suture, being associated with the rostral ridges ( Figure 8 View FIGURE 8 ); (3) the maxilla-maxilla contact, in the palatal surface, of M. latrubessei extends from the third to the sixth maxillary alveoli, while in M. niger (including MCNC 243), despite the intraspecific variability and asymmetrical development in some pathological specimens, this contact keeps restricted between the second and the fourth maxillary alveoli, or even less (e.g., in some specimens, this contact extends from the third to the fifth maxillary alveoli); (4) the anterior process of the ectopterygoid in M. latrubessei is inclined medially, stretching for one maxillary alveolus (the twelfth), while for M. niger (including MCNC 243), the anterior process is straighter and extends by two or three alveoli (reaching the twelfth/thirteenth maxillary alveoli); and, (5) despite the fact that M. latrubessei and M. niger exhibit two rostral ridges in the rostrum, it is important to mention that in M. latrubessei the rostral ridge 2 is developed and well-marked, being anteroposteriorly inclined, and completely composed by the maxilla without contacting the rostral ridge 1, while in M. niger (including MCNC 243), the rostral ridge 2 is located over or very close to the premaxilla-maxilla suture, extending from the dorsal surface until reaching the ventral margin of the aforementioned suture, being this ridge connected with the rostral ridge 2 ( Figure 8 View FIGURE 8 ).
All studied specimens (see Table 1) present a similar pattern of ornamentation, with irregular, but generally round and well-marked pits along the entirely dorsal surface of the rostrum. However, only in M. latrubessei this bone ornamentation does not reach the margins of the external nares ( Figure 2 View FIGURE 2 ). Though fragmented, the preserved portion of the external nares in M. latrubessei enables to infer a circular or keyhole-shaped format, as stated earlier ( Figure 2 View FIGURE 2 ); this condition (character83[0]) is shared with all other studied specimens (see Table 1).
The incisive foramen is only partially preserved in M. latrubessei ( Figure 3 View FIGURE 3 ), but since the lateral margin is well-preserved, based on the maximum curvature, it is possible to stipulate the anterior and posterior limits of the incisive foramen. In this scenario, the incisive foramen occupies less than half of the premaxillae maximum length (88[0]). The same condition is seen in other studied specimens, with exception of Caiman brevirostris and Caiman wannlangstoni Salas-Gismondi et al. 2015 , which do not have this portion preserved. Regarding the location of the incisive foramen, in relation to the premaxillary toothrow, it is inconclusive to assert that in M. latrubessei this foramen protrudes between the first premaxillary alveoli, as can be seen in the fossil specimen UCMP 39978 and in M. niger , as well as in Caiman species.
The anterior limit of the suborbital fenestra in M. latrubessei is located between the eighth and ninth maxillary alveoli, similar to observed in Caiman crocodilus (Linnaeus 1758) , Caiman latirostris (Daudin 1802) , Caiman yacare (Daudin 1802) , M. niger and UCMP 39978. As it was already mentioned, the suborbital fenestra in MCNC 243 (former M. fisheri ) reaches the space between the ninth and the tenth alveoli. The lateral margin of the suborbital fenestra comprised by the maxillary process is linear in all studied specimens (111[0]; Table 1), with exception of Caiman brevirostris and Caiman gasparinae Bona & Carabajal 2013, which do not preserve this portion.
The premaxilla exhibits, in dorsal view, two symmetrical posterior wedge-shaped processes that extend through three maxillary alveoli in Caiman brevirostris , C. crocodilus , C. latirostris , C. yacare , M. niger (including MCNC 243), M. latrubessei and UCMP 39978, in contrast to C. gasparinae , in which this projection does not surpass posteriorly any of that maxillary alveoli, being almost lateromedially straight. In ventral view, all specimens studied have the posterior premaxillary process restricted to the level of the first maxillary alveoli. Both conditions are unknown in C. wannlangstoni .
The nasal bones contact the posterior border of the external nares, but do not bisect it (82[1]), as seen in Caiman brevirostris , C. crocodilus , C. latirostris , C. wannlangstoni , C. yacare , and M. niger (at least some specimens, including MCNC 243), contrasting with the observed in C. gasparinae and M. latrubessei , in which the nasals are posteriorly retracted (at least externally) from the external nares, with the premaxillae contacting laterally in the medial line of the dorsal surface (82[2]). This condition is unknown in the UCMP 39978 specimen.
All studied specimens ( Table 1), with exception of Caiman wannlangstoni (that does not have this portion preserved), have the fourth dentary tooth leaving an occlusion pit in the format of a deep excavation, located between the last premaxillary and first maxillary alveoli, in the contact between these two bones. A similar condition is observed in M. latrubessei ( Figure 3 View FIGURE 3 ).
The anterior extension of the wedge-shaped ectopterygoid process lateral to the toothrow is quite variable in Caimaninae species. In M. latrubessei this process is inclined medially and extends for only one maxillary alveolus, while in M. niger (unknown in MCNC 243), the process is straighter and extends along the two or three most posterior maxillary alveoli. There are some species with an anterior process of the ectopterygoid elongated, that extends for three maxillary alveoli, such as Caiman latirostris , C. wannlangstoni and UCMP 39978, or even for four maxillary alveoli, such as in C. crocodylus and C. yacare . This condition is unknown in C. brevirostris and C. gasparinae .
The species that do not exhibit an exposed vomer, such as Caiman crocodilus , C. latirostris , C. yacare and the specimen UCMP 39978, have the maxilla-maxilla contact, in ventral view, extending for six maxillary alveoli. The only exception is C. brevirostris , where this contact extends for four maxillary alveoli. Among the species with an exposed vomer, M. niger (including MCNC 243) present the contact that extends for three maxillary alveoli while in M. latrubessei the maxilla-maxilla medial contact extends for four maxillary alveoli ( Figure 3 View FIGURE 3 ). This condition is unknown in C. gasparinae and C. wannlangstoni .
Considering mature specimens, the canthi rostralii is absent or very modest (96[0]) in Caiman crocodilus , C. gasparinae and C. yacare , contrasting with the very prominent condition (96[1]) observed in C. brevirostris , C. latirostris , C. wannlangstoni , M. niger , M. latrubessei and UCMP 39978 specimen. Associated to the well-developed canthi rostralii, there is the rostral ridge 1. The development of the rostral ridge 1 indicates prominent canthi rostralii in M. latrubessei . The rostral ridge 2 is developed at maturity only in M. niger and M. latrubessei , being not well-developed in the remaining studied species (see Table 1).
Remarks. Some comparisons use the relative position of the structure in reference to the maxillary alveoli. The number of maxillary alveoli in Caimaninae seems to have very few to null intraspecific variation ( Bona et al. 2017b), serving as a basis for other structure comparisons. However, as M. latrubessei and M. niger have different number of alveoli in the maxilla, those comparisons could be equivocated. This alveoli variation between Melanosuchus species could be result of two processes equally possible: (1) either the thirteenth alveoli was reduced and disappeared in M. latrubessei ; or (2) it emerged and was fixed among M. niger specimens. Also, all references were based on the middle to anterior alveoli that seems to have a homologue correspondence between these species, based on their morphology. Therefore, this different alveolar number in maxilla cannot be considered a problem with the references provided by the other structures. The only exception for these morphological references is the anterior extension of the ectopterygoid, where in M. niger extends for two maxillary alveoli, while in M. latrubessei , it extends for one and this variation seems to be biologically dependent with the different alveolar count that is varying in the posterior region of maxilla. Therefore, it is not possible to assure that this difference in alveolar count is the result of a well-extended anterior process of the ectopterygoid in M. niger or due to the absence of an alveolus in M. latrubessei .
The rostral ridge 2 varies in size and robustness among the studied specimens of M. niger (including MCNC 243), and this variation is strongly correlated to the maturity of the specimens, with the ridge being more robust and developed in large and adult animals ( Figure 8 View FIGURE 8 ). However, in none of the observed skulls the rostral ridge 2 presents significant variation in position, being located over or very close to the premaxilla-maxilla suture in all specimens (e.g. Figure 8 View FIGURE 8 ). Additionally, the rostral ridge 2 in M. latrubessei is completely isolated from the rostral ridge 1, which is opposed to the feature observed in all studied M. niger ( Table 1). Therefore, the present work considers that the rostral ridges 2 in M. latrubessei and M. niger are homologous structures, even located in different portions of the rostrum. Such differences in position and morphology are related to interspecific variation.
The general prismatic morphology of the vomer among Melanosuchus specimens (e.g. Brochu 1999; Vieira et al. 2016) is quite variable, going from a more circular to a more angular prismatic shape. This morphological variation is reflected in the morphology of the premaxilla and maxilla, but in all studied specimens the pattern described here ( Figure 7 View FIGURE 7 ) was observed. In Melanosuchus , the morphology and position of the vomer has some similarities with Squamata species (e.g. Jollie 1960; Avery & Tanner 1971; Gauthier et al. 2012), however, there is no available discussion or literature about the Melanosuchus vomer. Thus, further studies are necessary for a better understanding regarding the embryology, ontogeny and function of the structure.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |