
Formica foreli Emery, 1909
|
publication ID |
https://doi.org/10.5281/zenodo.5392741 |
|
persistent identifier |
https://treatment.plazi.org/id/03BB87B2-FF98-F14F-4D27-FB43FF69FE86 |
|
treatment provided by |
Marcus |
|
scientific name |
Formica foreli Emery, 1909 |
| status |
|
Formica exsecta pressilabris var. foreli Emery, 1909 TYPE LOCALITY. — Vaux near Morges, Switzerland, leg. Forel.
TYPE MATERIAL. — Holotype worker ( MCSN) [investigated].
Formica naefi Kutter, 1957 . New synonym.
TYPE LOCALITY. — Scuol-Pradella (Bain Jonnair), Switzerland
TYPE MATERIAL. — Syntypes 13 males, 5 queens,> 80 workers ( MZ) [investigated].
Formica tamarae Dlussky, 1964 . New synonym.
TYPE LOCALITY. — Omaló, Caucasus, leg. Zhizhilashvili.
TYPE MATERIAL. — Paratype and topotypical samples leg. Zhizhilashvili & Seifert [investigated].
25
5.2 4.4 3.6 2.8 2.0 1.2 0.4 0.4 1.2 2.0 2.8 3.6 4.4 5.2 – – – – – – – discriminant D(4)
Formica goesswaldi Kutter, 1967 . New synonym.
TYPE LOCALITY. — Soglio in Val Bregaglia, Switzerland. TYPE MATERIAL. — Syntypes 20 males, 3 queens, 56 workers ( MZ) [investigated].
GEOGRAPHIC ORIGIN OF THE MATERIAL STUDIED. — The numerically evaluated 421 specimens ( 330 workers, 69 queens, 22 males) came from Sweden 42, Germany 190, Czechia 8, Slovakia 7, Switzerland 124, Austria 4, Spain 8, Turkey 16, and Caucasus 22. Total number of specimens seen> 600.
DESCRIPTION
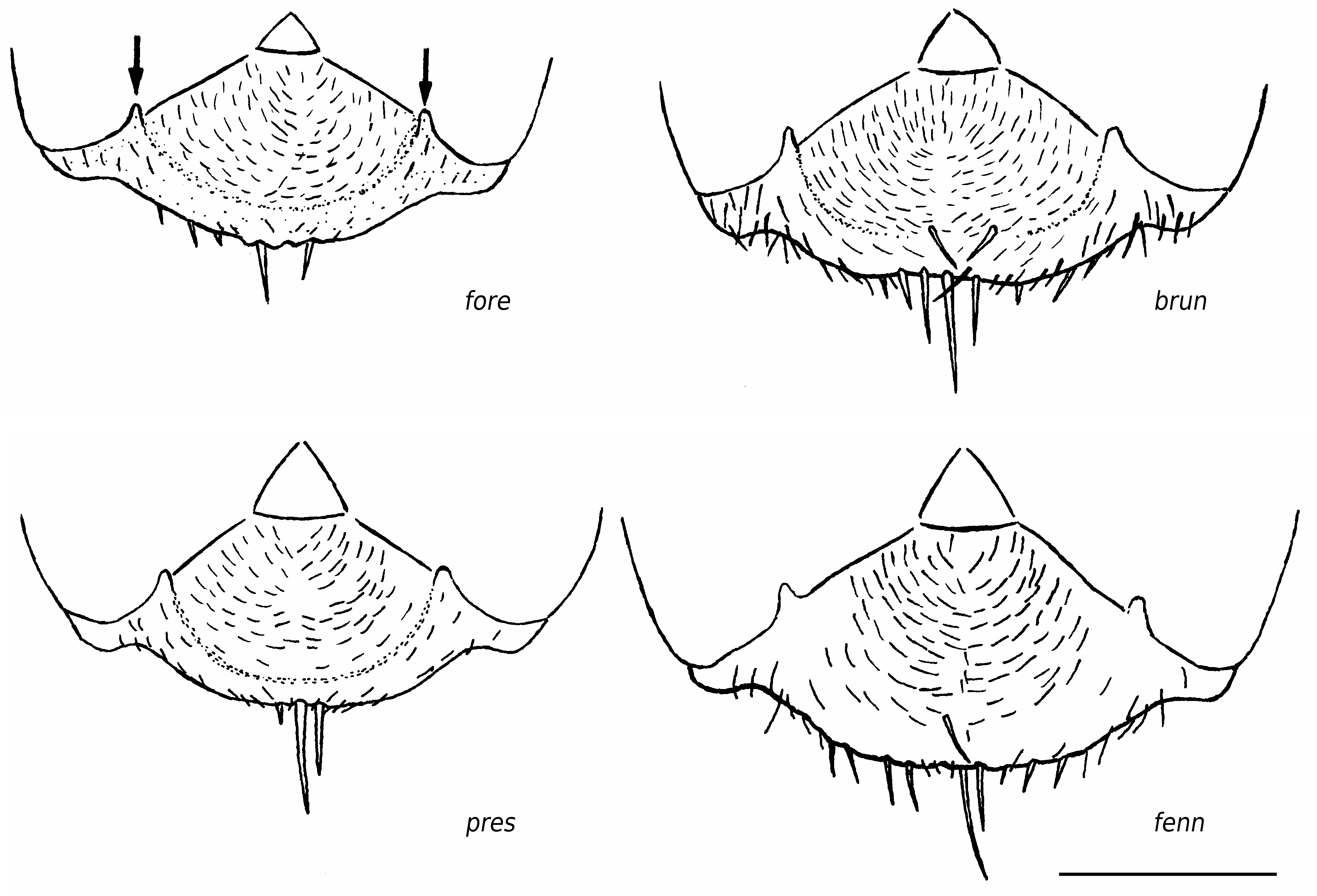
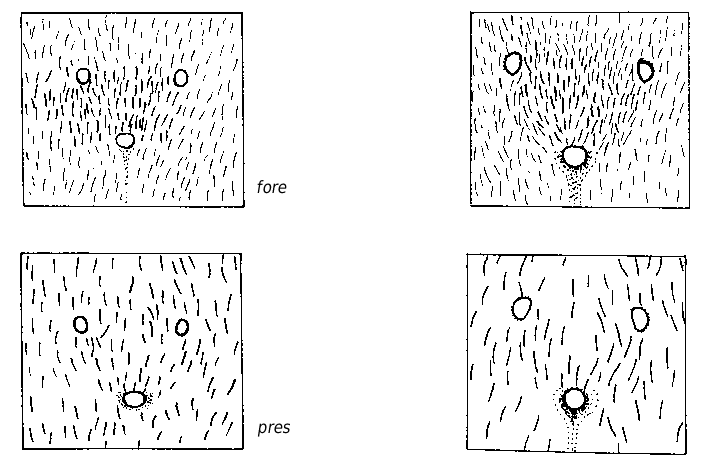
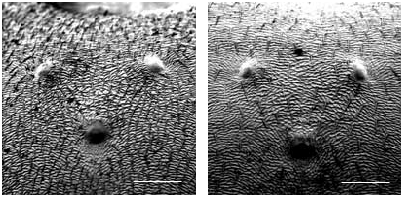
Worker ( Figs 3 View FIG ; 4 View FIG ; 7 View FIG ; 9 View FIG ; 13 View FIG ; 16)
Rather small (CL 1288 ± 79, 1013-1472; CW 1219 ± 77, 981-1371). Head moderately elongated (CL/CW 1.057 ± 0.020, 0.991-1.120). Scape rather long (SL/CL 1.008 ± 0.023, 0.937- 1.083). Clypeal setae restricted to anterior margin ( Figs 3 View FIG ; 16), a small second level seta is in 7% of specimens present (ClySet 1.08 ± 0.27, 1-2). Clypeus lateral of the tentorial pit level only very exceptionally with single pubescence hairs surpassing the anterior margin by more than 10 µm (ClyPub 0.04 ± 0.23, 0-2.0, Fig. 16). Lateral semierect setae in the ocellar triangle always absent (OceSet 0%). Eye hairs fully absent or few minute hairs present (EyeHL 5.0 ± 2.1, 0-11; Fig. 9 View FIG ). Pubescence hairs in the occellar triangle dense (sqrtPDF 4.44 ± 0.44, 3.36-6.16; Figs 4 View FIG ; 13 View FIG ). Craniad profile of forecoxae without setae (nCOXA 0.0 ± 0.0). Lateral metapleuron and ventrolateral propodeum without standing setae (nMET 0.0 ± 0.0). Outer edge of the hind tibial flexor side with few semierect first order setae, second order setae absent (nHTFL 2.67 ± 1.16, 0-6.0). Erect setae on gaster often beginning on posterior margin of fourth tergite (TERG 3.88 ± 0.40, 3-5). Pubescence on first gaster tergite variable, but usually relatively dense (sqrtPDG 6.09 ± 0.44, 4.55-7.34).
Queen ( Fig. 6 View FIG )
Significantly larger than pressilabris (CL 1341 ± 38, 1279-1464; CW 1362 ± 38, 1286-
4.8 4.0 3.2 2.4 1.6 0.8 0.0 0.8 1.6 2.4 3.2 4.0 4.8 5.6 6.4 7.2
– – – – – –
discriminant D(5)
1479; ML 2227 ± 57, 2107-2415). Head proportions of average Coptoformica type (CL/CW 0.985 ± 0.022, 0.934-1.042), scape longer than in pressilabris (SL/CL 0.910 ± 0.023, 0.857- 0.962). Clypeal setae restricted to anterior margin, second level setae only exceptionally present (ClySet 1.01 ± 0.12, 1-2). Clypeus lateral of the tentorial pit level without pubescence hairs surpassing the anterior margin by more than 10 µm (ClyPub 0.0 ± 0). Erect setae in the ocellar triangle absent. Eye hairs absent or short (EyeHL 6.1 ± 3.6, 0-21). Pubescence in the occellar triangle very dense (sqrtPDF 4.03 ± 0.32, 3.36- 4.77). Occipital corners of head with fully appressed pubescence (OccHD 0.0 ± 0.0). Dorsal head surface variable, on average less shining than in pressilabris (GLANZ 1.87 ± 0.35, 1.0-2.5). Craniad profile of forecoxae without setae (nCOXA 0.0 ± 0.0). Dorsal mesosonotum and scutellum without standing setae and only appressed pubescence (MnHL 0.0 ± 0.0). Outer edge of the hind tibial flexor side with very few suberect to subdecumbent first order setae, second order setae absent (nHTFL 1.23 ± 0.62 0- 2.5). Erect setae on gaster tergites usually beginning at the posterior margins of fourth to fifth tergite (TERG 4.65 ± 0.56, 2-5). Pubescence on first gaster tergite usually dense (sqrtPDG 5.42 ± 0.54, 4.54-6.68). Whole body less shining than in pressilabris .
TAXONOMIC COMMENTS AND
DIFFERENTIAL DIAGNOSIS
The holotype specimen of Formica foreli was collected by Forel at Vaux near Morges/ Switzerland and is still present in MCSN Genova. Bondroit (1918) stated that queens of foreli are different from those of pressilabris by the dull body surface, the slightly larger size, and the more dense gaster pubescence, i.e. his differential diagnosis is in agreement with the species conception presented below as it is in agreement with the investigation of Emery’s type worker.
Agosti (1989) synonymised Formica foreli with pressilabris . One reason for his decision were three puzzling nest samples collected by Malicky near Montana/ Switzerland which are stored in MZ. These samples represent mixed nests of pressilabris and bruni , which contain a large fraction of true hybrids bruni x pressilabris . A detailed argumenta- tion leading to this conclusion was presented elsewhere (Seifert 1999). The second reason for Agosti’s synonymisation were Swedish foreli and pressilabris samples with ambiguous gaster pubescence. Such samples are not rare in S Sweden but, based upon character combinations, they can be allocated to either foreli or pressilabris in a similar way as in other geographic regions (see below).
Significant differences in external morphology, zoogeography, and habitat selection give convincing evidence for a heterospecifity of foreli and pressilabris . The worker of foreli differs by a much more dense frontal pubescence, a significantly denser tergite pubescence, the more caudal position of tergite setae, and a longer scape. In these diagnostic characters, the holotype specimen of foreli fits very well to the population average with SL/CL 1.025, Terg 4.0, sqrtPDF 4.59 and sqrtPDG 6.07. A discriminant score D(4) calculat- ed with the characters sqrtPDF, sqrtPDG, TERG, and SL/CS provides a clear separation of both species in 181 investigated W Palaearctic nest samples ( Table 2; Fig. 6 View FIG ). The D(4) of the type series of goesswaldi and naefi , of the holotype of foreli , and of the topotypical series of tamarae are fully within the cluster defined here for Formica foreli while the type series of pressilabris and rufomaculata are clearly within the pressilabris cluster ( Fig. 6 View FIG ). Queens of foreli and pressilabris are separable on the individual level by the characters sqrtPDF, sqrtPDG, SL/CL, ML, and GLANZ. A discriminant score D(5) calculated with these characters provides a clear distinction ( Table 3). All available type queens of goesswaldi and naefi , and one paratype queen of tamarae have D(5) values very distant from the pressilabris cluster ( Fig. 7 View FIG ).
Further evidence for a heterospecifity of foreli and pressilabris is provided by zoogeography and habitat selection. Formica pressilabris represents a boreo-alpine-continental and foreli a submediterranean species with northern range expansion. Zones of geographic overlap are rather small and known from S Sweden, W Slovakia, and the W Alps. The external morphology of foreli and pressilabris samples from within and from outside the overlap zones does not differ significantly indicating that both species do not hybridise in detectable frequencies. The vertical distibution in the W Alps is very weakly overlapping. F. foreli usually occurs below 1600 m and is preferentially a species of warm valleys on spots with very xerothermous mediterranean floral elements. F. pressilabris , in contrast, mainly occurs in altitudes of 1750-2250 m and is a species of thermophilic mountain meadows and pastures with elements of boreo-alpine flora. Vaux near Morges, the type locality of foreli , is situated at 552 m. This fact gives additional evidence that foreli cannot be referred to pressilabris .
The synonymisation of Formica goesswaldi , naefi , and tamarae with foreli needs explanation. The investigated topotypical material of goesswaldi from Soglio and Vicosoprano/Val Bregaglia comprised five nest samples with 23 workers and three queens. The population of Formica naefi was studied in 14 nest samples with 57 workers and in 13 queens from the terra typica in the Unterengadin (Pradella/Scuol, Bain Jonnair/Scuol, Ramosch) and included all available types. Formica tamarae was studied in five nest samples with 21 workers and one paratype queen from the type locality Omaló/Caucasus. The data of Table 4 show only weak differences between the considered populations in the 13 evaluated characters which cannot even justify a subspecies rank. Formica tamarae is fully identical with the Central European foreli except for the slighly lower TERG data. The only difference of goesswaldi is the larger CL/CW that is, however, within the known range of variation in foreli . Formica naefi deviates by a significantly lower CL/CW and sqrtPDF but again these data do not exceed the range known in foreli .
The situation in the queens is similar ( Table 5). In none of the 12 characters evaluated is a difference suggesting a heterospecific identity of the four taxa. The elongation of the head in goesswaldi considered as diagnostic by Kutter (1977) is within the range known for foreli as it is the broadening of head in naefi . Differences in mesosoma shape, pigmentation, and scale shape, which were considered as diagnostic by Kutter, are in fact invalidated by the high intraspecific variation of these characters. In one character, however, the topotypical queens of naefi show an unusual deviation: they have a significantly larger eye size (p <0.001). The ratio (EL + EW)/(CL + CW) is 0.293 ± 0.007 [ 0.280 -0.306] in 13 naefi queens from the Unterengadin and is 0.282 ± 0.007 [ 0.267 -0.296] in 37 foreli queens from outside this region. The weak deviations of this relatively isolated population in the Swiss Alps (which is actually threatend by extinction!) are interpreted here as consequence of bottle-necking events in the past and not as indication for a heterospecific status.
BIOLOGY AND DISTRIBUTION
Geographic range
Formica foreli represents a submediterranean species with northern range expansion during warmer climatic periods and subsequent splitting into isolated populations after regional extinction. It is known from N Spain, N Italy, Anatolia, the Caucasus, the W Alps, S Moravia, and W Slovakia. Reliable records from the German countries refer to an isolated population in Thüringen, two local populations in N Sachsen, larger populations in Brandenburg and the southern Mecklenburg-Vorpommern, and two local populations in Schleswig-Holstein. An isolated northern population exists in NE Zealand/ Denmark and Skåne /S Sweden with nine sites known. The population in the W Alps is restricted to valleys with xerothermous local climate in altitudes of 1181 ± 348 m (552-1780), (n = 10). In more southern latitudes ( Anatolia) it goes up to 2200 m.
Habitat selection
F. foreli is a species of open oligothrophic and xerothermous grasslands mainly on sand, but also on limestone and other geological outcrop. The different zoogeography and much more expressed xerothermy of foreli compared to pressilabris is reflected by plant sociology. In all habitats studied at least some indicator plants for strong xerothermy such as Festuca cinerea , Festuca ovina, Euphorbia cyparissias, Hieracium pilosella , Echium vulgare , or Eryngium campestre were present. Very strong polycalic colonies in Brandenburg were observed to extend their nesting area into mesophilic grassland and light woodland but became extinct in the latter after full closure of the canopy.
Status as threatened species
Formica foreli is one of the most endangered species of the group. In Switzerland it is threatened by extinction. In Germany it has the same status, though the populations in Brandenburg and Mecklenburg-Vorpommern seem less endangered. The habitat destruction is mainly caused by intensive use of mineral fertilisers and liquid manure, high athmospheric nitrogen immission, decline of sheep pasturing and traditional cutmeadow management, intensifying of cattle pasturing, and afforestation programs.
Colony foundation
Four pins with Formica foreli workers from the type series of Formica naefi (labelled “Bain Jonnair, 10.7.54, Kutter-/”) carry four workers of Formica lemani and one worker of Formica lusatica . The equal labelling should indicate a nest sample but the mode of sampling is not explicitely stated. Observations of Kutter (1957, 1969) in an laboratory arena with nests of lemani and foreli (his naefi ) showed an eudulotic behaviour of foreli with subsequent killing of the lemani queen. It seems possible that eudulotic behaviour can also occur in nature (callows at least can be taken) and that mixed nests must not necessarily indicate a preceeding socially parasitic colony foundation by a single foreli queen. Habitat selection and zoogeography of foreli should exclude lemani as primary host species except for few alpine populations. The main host species in the whole geographic range should be either fusca , cunicularia , or rufibarbis .
Nest construction
There is no difference from the normal Coptoformica type. The mounds do not reach the size known for exsecta and their diameter is usually < 50 cm. The diameter of the subteranean part of the largest nest was 80 cm. On very xerothermous spots of sand dunes or sandy moraines, where the field layer is very sparse, nests may entirely lack any vegetable cover and are only circumscribed by simple entrances in the soil surface. Nests in an intensive sheep pasture near Carwitz/ Germany had constucted their subterranean galleries within the solidified root bale of vegetation to increase the resistance against trampling.
Demography of nests and colonies
No reliable information on the nest population is available but figures similar to bruni are expected. The tendency towards polygyny and polycaly is very expressed. The largest polycalic colonies in Brandenburg comprised 100 nests/ 2500 m 2 and 78 nests/ 1200 m 2. Polycalic colonies have a sex ratio> 1 and produce mainly micraners.
Swarming
Alates were observed in the nest 14.2 July ± 24.9 d (12 June-24 Aug, n = 10) which is somewhat earlier than in pressilabris . Modalities and timing of swarming are unknown.
Intra- and interspecific behaviour
F. foreli is very aggressive and territorial against other territorial Formica species and Lasius .
Food sources
Workers of a large polycalic colony at the margin of a light forest in Brandenburg visited in large files Aphidae on Pinus silvestris and Betula pendula trees.



| MCSN |
Museo Civico di Storia Naturale, Verona |
| MZ |
Museum of the Earth, Polish Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |