
Cordagalma bimaculatum, P. R. Pugh, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4095.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:690FFEBE-F71B-4EFD-865A-944D81A12897 |
|
DOI |
https://doi.org/10.5281/zenodo.6087821 |
|
persistent identifier |
https://treatment.plazi.org/id/03BC87D0-FF85-FFFA-FF6E-FABBF806D03F |
|
treatment provided by |
Plazi |
|
scientific name |
Cordagalma bimaculatum |
| status |
sp. nov. |
Cordagalma bimaculatum sp. nov.
Cordagalma cordiforme Mills et al., 1996 , p. 154. Cordagalma ordinata Mapstone, 2009 , Fig. 22 View FIGURE 22. A D.? Cordagalma cordiforme Mackie, 1985
Diagnosis. Nectophores heart-shaped, with a distinctive patch of large ectodermal cells at the apex of each axial wing. Broad axial wings with deep narrow incision between them. Usually two pairs of patches of ectodermal cells on lower, lateral parts of velum. Bracts roundly rhomboidal in shape, with a transverse ridge on the upper side dividing off a distal facet. Small nematocysts can line the margins of the facet, and also form a median line dividing the distal facet. A patch of ectodermal cells present on each side of the facet. Bracteal canal extends to half the length of the bract and, in younger ones, has a small swollen extension into the mesogloea. Young tentilla without long distal cnidocils. Palpon attached laterally.
Material examined. Two specimens collected by the Johnson-Sea-Link ( JSL) submersibles:
JSL II Dive 1405 39°51.6’N, 70°22.5’W 1-Sep-1988 depth 747 m GoogleMaps JSL I Dive 2952 36°02.9’N, 02°48.2’W 17-Apr-1991 depth 726 m GoogleMaps
Unfortunately the nectosome of the JSL II Dive 1405 specimen became detached during collection and was lost.
A further six specimens were collected by various MBARI submersibles, namely: Ventana Dive 209 36°42.69'N 122°01.15'W 22-Jan-1991 Depth 422 m GoogleMaps Ventana Dive 3005 36°41.85'N 122°03.94'W 15-May-2007 Depth 372 m GoogleMaps Tiburon Dive 843 35°29.87'N 23°51.91'W 08-Apr-2005 Depth 587m GoogleMaps Tiburon Dive 1043 36°42.15'N 122°34.49'W 4-Oct-2006 Depth 400 m GoogleMaps Ricketts Dive 419 36°36.00'N 122°09.03'W 28-Sep-2012 Depth 449 m GoogleMaps Ricketts Dive 500 36°36.00'N 122°09.00'W 15-Jul-2013 Depth 302 m GoogleMaps
Holotype: The specimen collected during the Johnson-Sea-Link I Dive 2952 has been designated the holotype, and will be deposited at the United States National Museum (Smithsonian Institution), Washington, DC.
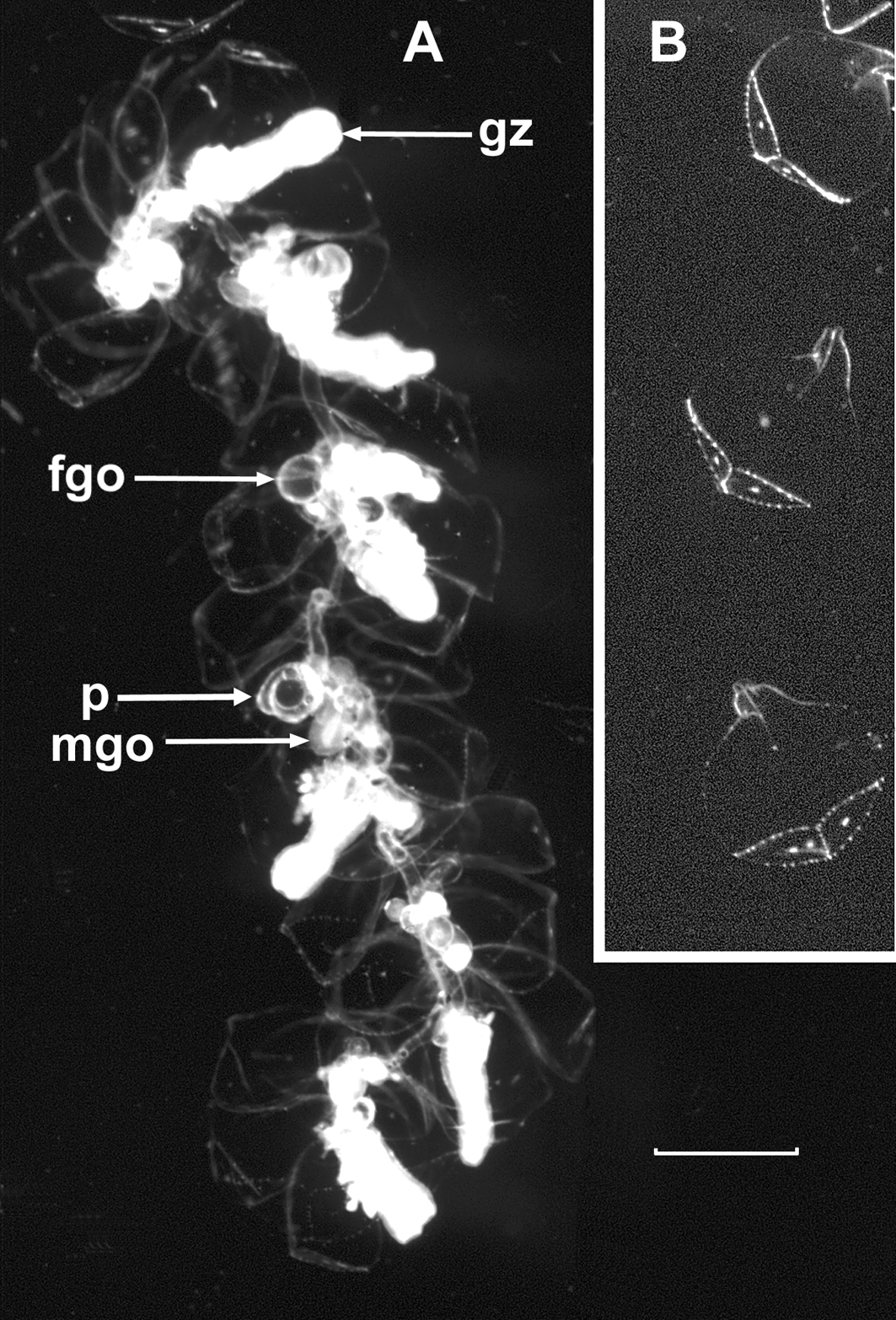
Description. The description of Cordagalma bimaculatum sp. nov. is based, for the most part, on the JSL specimens. However, there was some slight variability in the characters of certain zooids between the JSL and Monterey Bay material, to which attention will be drawn. A photograph of the preserved holotype specimen is shown in Figure 27 View FIGURE 27 . All the nectophores had become detached.
Pneumatophore: The pneumatophore ( Figure 27 View FIGURE 27 , pn) was distorted due to rupture of its basal wall caused by gas expansion as the specimen was brought to the surface. It measured approximately 0.55 x 0.25 mm and showed no signs of pigmentation, although there was a distinctive cap of cells apically.
Nectosome: The nectosome of the preserved type specimen ( Figure 27 View FIGURE 27 , n), despite the contracted state of the colony, was relatively long, and bore six nectophoral buds at its anterior end ( Figure 27 View FIGURE 27 , nb). The nectophores, as indicated by their attachment lamellae, were clearly attached to the ventral side of the nectosome.
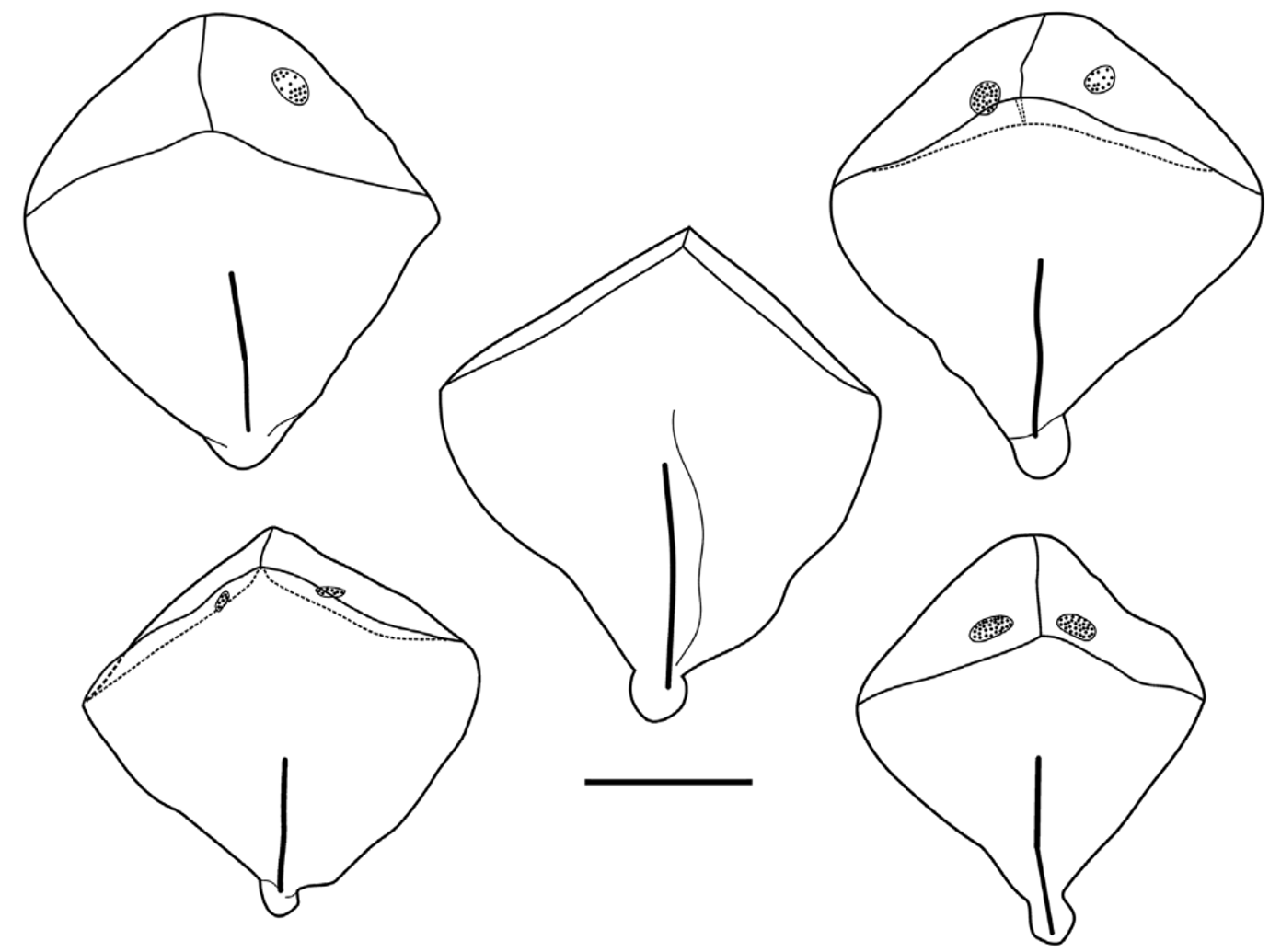
Nectophore: ( Figures 28 View FIGURE 28 , 29 View FIGURE 29 ). Despite the apparent length of the nectosome, only seven mature nectophores, which measured up to 3.8 mm in length and 3 mm in width in their preserved state, remained with the holotype specimen. These were heart-shaped, with a pronounced basal extension. The extent and shape of this basal extension was quite variable, possibly as a result of preservation; sometimes being pointed ( Figure 29 View FIGURE 29 A), and sometimes broadly rounded ( Figure 28 View FIGURE 28 ). The maximum number of nectophores found with any of the Monterey Bay specimens was 10 for the Tiburon Dive 843 specimen.
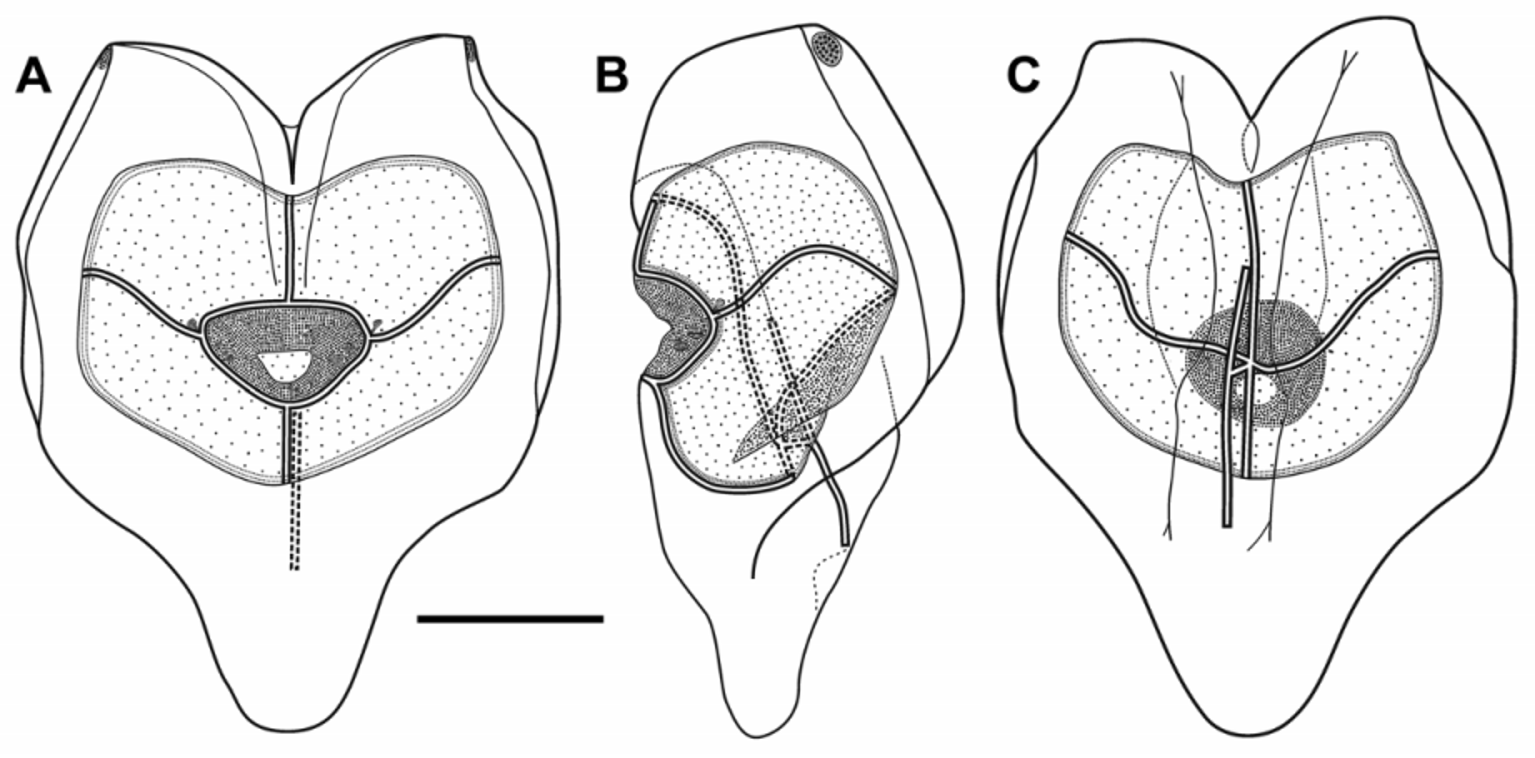
There was a deep median incision between the axial wings. On the mature nectophores, on the ostial side, a pair of weak ridges ran from the outer extremes of the axial wings toward the mid-line and continued toward the ostium, demarcating a shallow gutter between them, before petering out slightly above it. A very weak lateral ridge extended down the main body of the mature nectophore, on each side, but this could not be seen on the younger ones. Another pair of weak ridges, on the axial side of the nectophore, demarcated a relatively narrow hydroecial gutter between them, through which the nectosomal stem had once passed. At the upper end of the axial wings, just axial to the lateral ridge, there was a pair of small patches consisting of up to twenty large, ectodermal cells, probably photocytes, with darkly staining nuclei. Although characteristic these patches were unfortunately often very difficult to discern without staining, or had been abraded off in poorly preserved specimens such as the Ventana 209 one. They could be seen on only one of the four nectophores found with the Ventana Dive 3005 specimen. The more mature of these nectophores appeared to have an extensive basal process, but this probably was an illusion caused by the excessive contraction of the nectosac during preservation.
The ascending and descending mantle canals were of about equal length. The nectosac was also heart-shaped and occupied most of the main body of the nectophore. On reaching the nectosac the pedicular canal immediately gave rise to all four radial canals. The upper and lower canals ran straight to the ostial ring canal. The laterals looped up and over on to the lateral sides of the nectosac, before curving gently down and continuing to the ostial ring canal. This latter canal surrounded a large velum. In the region where the lateral radial canals joined the ostial ring canal, there was a club-shaped patch of distinctive ectodermal cells that extended out laterally for a short distance, although they could not always be discerned. In addition, on the lower lateral sides of the velum itself there were variably sized patches of cells, usually two on each side. On the younger nectophores this consisted of a single enlarged triangular patch.
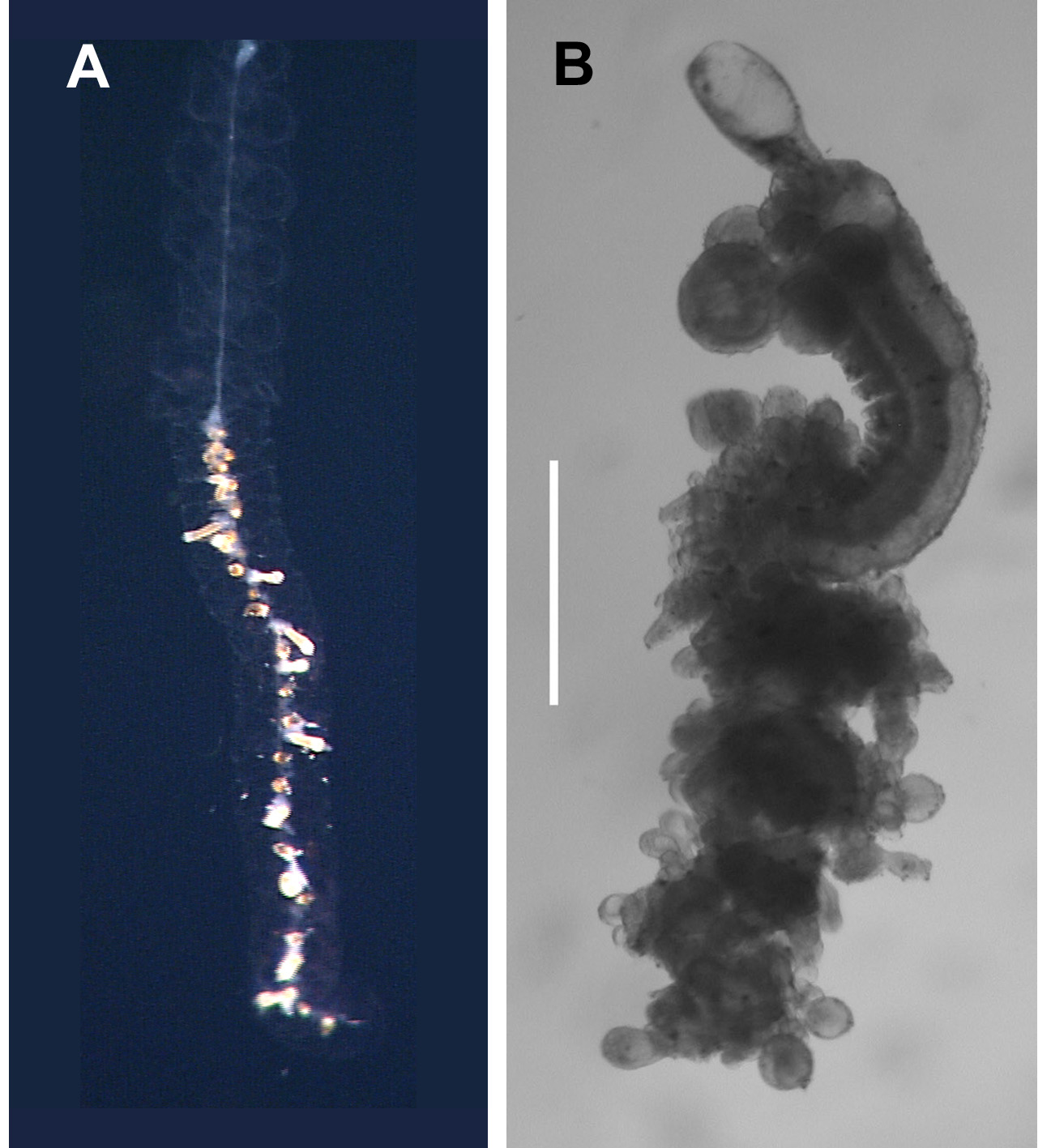
Siphosome: The highly contracted siphosome of the type specimen can be seen in Figure 27 View FIGURE 27 , and the slightly less crowded siphosome of the other specimen in Figure 30 View FIGURE 30 .
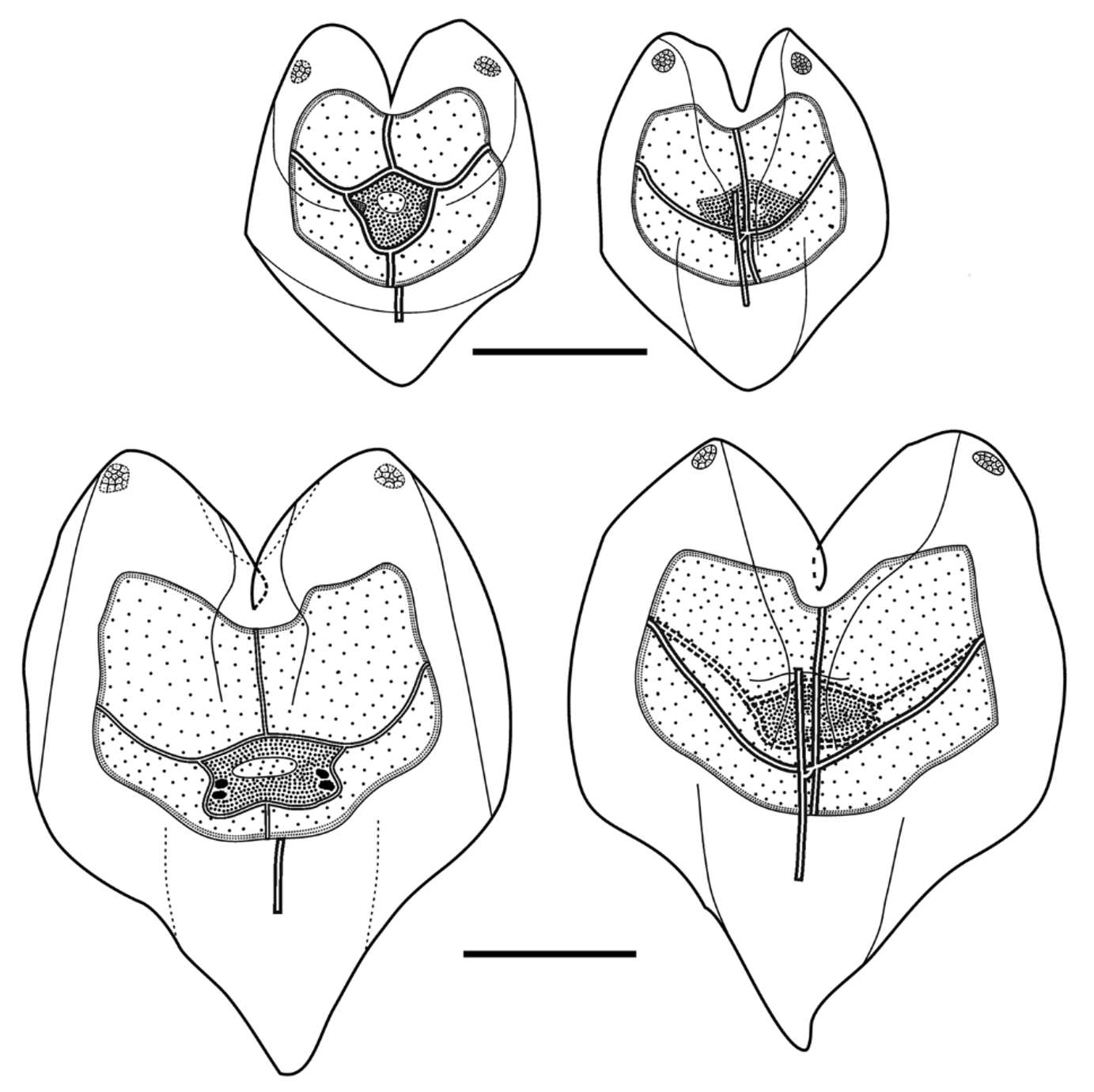
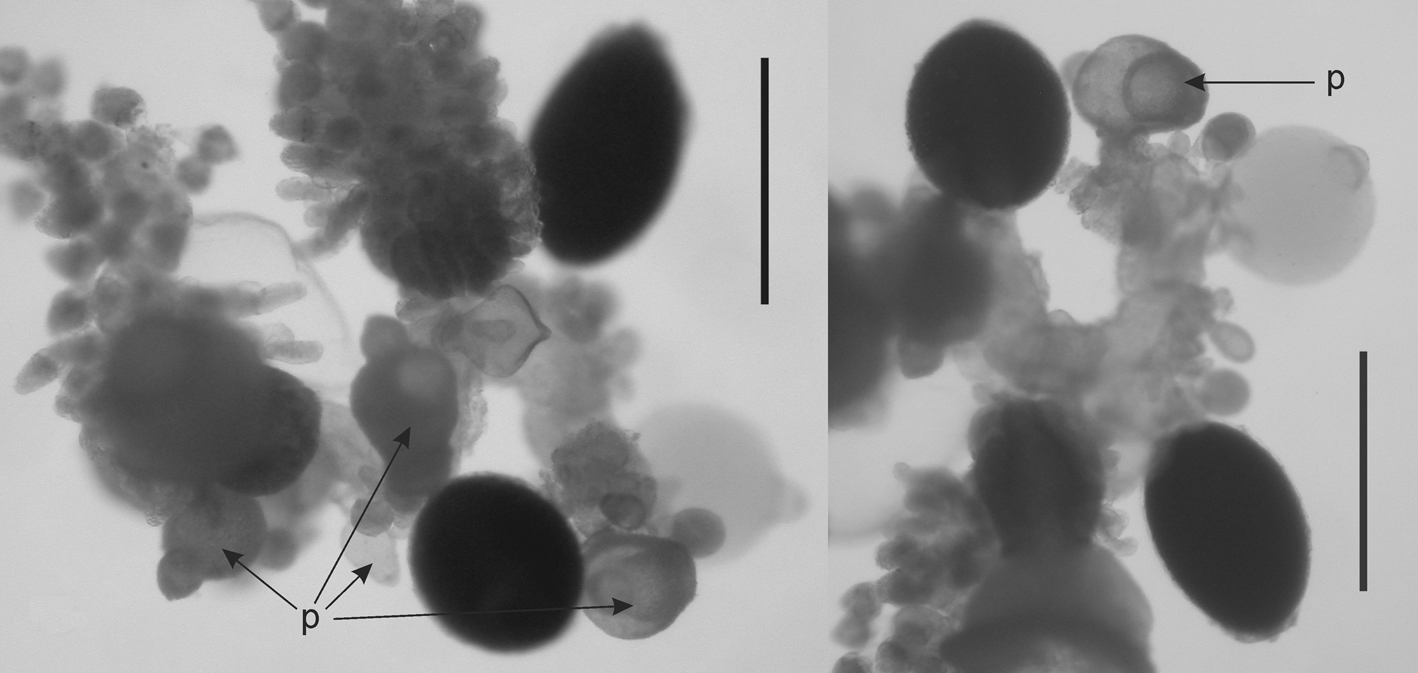
Bract: Numerous small bracts ( Figure 30 View FIGURE 30 B, 31–34), measuring up to c. 2.25 mm long and 2 mm wide, were present with both specimens. Those of the type specimen had a rounded roughly rhomboidal shape. They were slightly concave, centrally, on the lower side, and distinctly convex on the upper one. Toward the distal end a weak transverse ridge demarcated a rapid decrease in the thickness of the bract on the upper side. The whole surface of the preserved bract had a papillose appearance, particularly on the distal facet. The two sides of that facet joined, in the mid-line, forming an angle of c. 120°. On that facet there were two patches of large, distinctive ectodermal cells, which probably were sites of bioluminescence. Each patch usually consisted of 6–12 cells, but several had less. These patches were not always apparent on the younger bracts. In addition there was a median row of rounded cells, which probably were minute nematocysts, although this could not be ascertained with certainty. In life, these cells were also spaced out along all the edges of the distal facet, but after preservation many of these were abraded, and were less obvious in the JSL I Dive 2952 (holotype) specimen than the JSL II Dive 1405 one. The bracteal canal ran from the proximal end of the bract to approximately its mid-length.
For some of the specimens from Monterey Bay, the transverse ridge tended to be much more distinct and, like the younger bracts of the type specimen, it had a more pronounced V-shape. The bracteal canal, for all specimens, did not extend beyond half the length of the bract and greatly thickened in the younger bracts ( Figure 32 View FIGURE 32 ), and its distal end was swollen and inflected into the mesogloea. In the mature bract that swelling had disappeared. The canal usually originated above the proximal end of the bracts ( Figure 33 View FIGURE 33 ), although for some bracts there was a narrow extension to the proximal tip. The array of nematocysts around the edges of the distal facet, particularly on the upper transverse ridge and in the mid-line between the two facets, was clearly to be seen on the younger bracts. This was particularly noticeable on the very young bracts of the Ventana Dive 3995 specimen, which had nematocysts inserted without intervals demarcating the upper and median borders of the distal facet.
Amongst the Monterey Bay specimens just one, from Tiburon Dive 843, had only bracts resembling those found with the JSL specimens. Three others had only bracts of the type found with the Tiburon Dive 1043 specimen ( Figure 33 View FIGURE 33 ), while the specimen from Doc Ricketts Dive 419 had a combination of both types of bract in approximately equal numbers ( Figure 34 View FIGURE 34 ). With so few specimens it is impossible to speculate on any possibly reasons why these variations in the bract type occur.
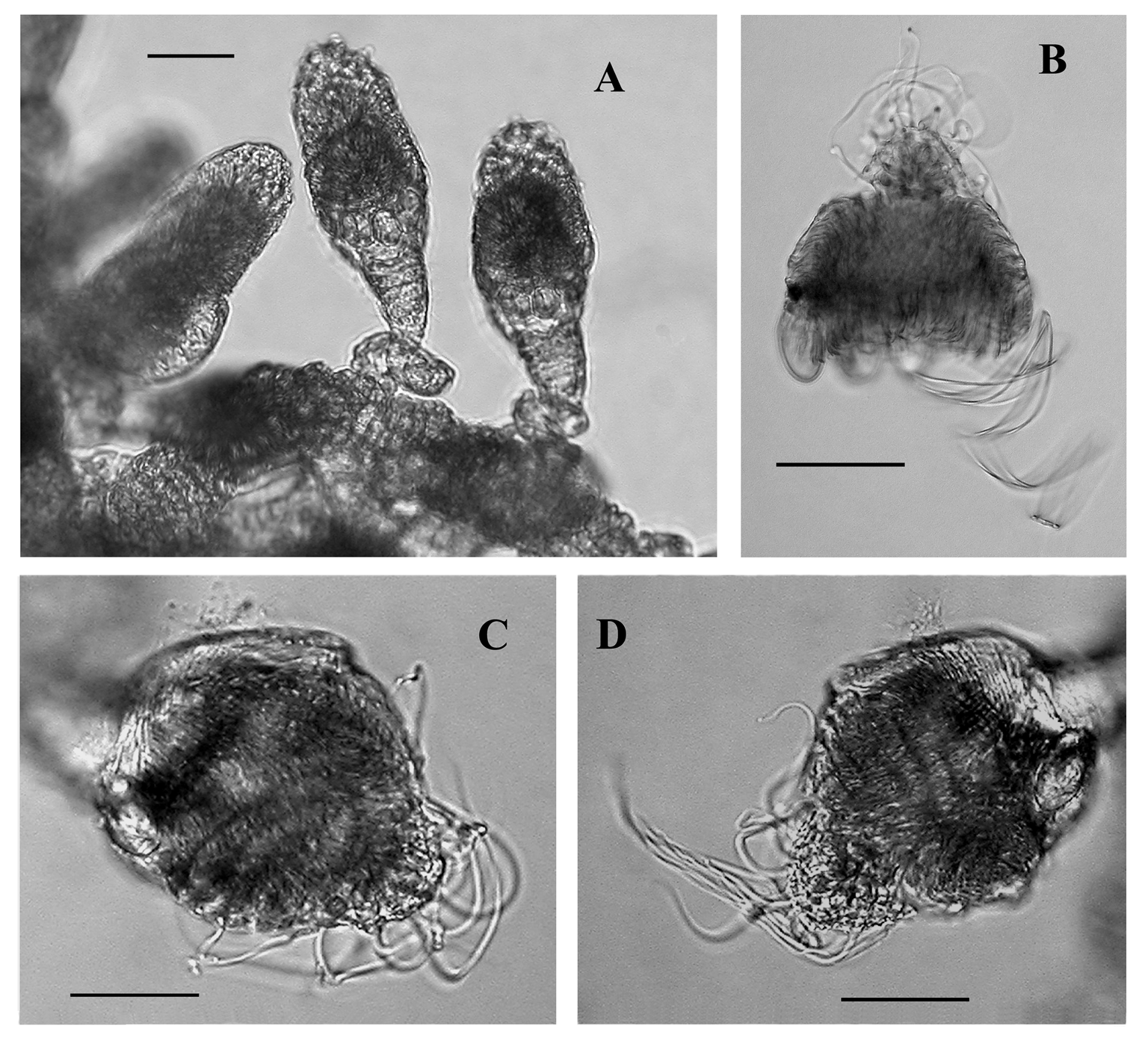
Gastrozooid: The small gastrozooids measured up to 1.5 mm in length, and had a distinct basigaster measuring up to 0.3 mm in length ( Figures 35–36 View FIGURE 35 View FIGURE 36 ), surmounted by an inflated stomach and a more or less extended proboscis. Dark brown droplets of oil were occasionally found at the base of the gastrozooids. The gastrozooids were, as usual, of very variable shape, not least those with the Ventana Dive 3005 specimen where there was a very large basigaster, a short narrow stomach that looked as if it formed the stalk for a toadstool-like proboscis region.
Tentilla: The proximal end of the tentacle was always highly contracted and it was impossible to make out the early stages in the development of the tentilla. The youngest tentilla observed ( Figure 37 View FIGURE 37 A) were in the process of developing the cnidoband, and the cnidocils at their distal end were only rudimentary. The mature tentilla ( Figure 37 View FIGURE 37 C, D) were very similar to the type of variant tentillum, without a rostrum, of Cordagalma ordinatum that Carré (1968) mentioned and figured in his Plate III, figure 7. Indeed the cnidobands of both included five proximal, elliptical stenoteles, but for C. bimaculatum sp. nov. they were about twice as large, measuring 30 x 15 µm, as opposed to 17.5 x 8.5 µm for Carré's specimens. These stenoteles were grouped together on one side of the cnidoband. The other nematocysts in the cnidoband were not examined in detail, but were presumed to be the numerous anisorhizas in the middle and distal section, with a few distal isorhizas. At the distal end of the cnidoband there were numerous long cnidocytes, which in life were probably straight, but in the preserved state often twisted and distorted. No rostrum was present.
One curious feature of the tentilla was that if pressure was applied to them the basal part of the cnidoband, with the exception of the stenoteles, became detached from the rest and was found ( Figure 36 View FIGURE 36 B) to consist of a concertinaed, flattened tube that connected directly to the pedicle. Presumably, if fluid were to be pumped into it, it would expand to form a long straight tube. What part this would play in prey capture is uncertain, but such an arrangement has not previously been observed in other siphonophore species.
Palpon: The palpons ( Figures 38–39 View FIGURE 38 View FIGURE 39 ) were very small, up to 0.5 mm in length. They were roughly spherical in shape, with a more or less pronounced proboscis. For the larger palpons, at least, the short peduncle, which attached it to the stem, was placed laterally (see Figure 27 View FIGURE 27 ). The exact mode of attachment of the developing palpons was difficult to discern. The palpons were without a palpacle. No nematocysts were found within the palpons. The palpons frequently contained golden or dark brown oil droplets of variable size; some occupying about a third of the interior. They were usually found laterally on the side opposite to the proboscis. Several of these were also present within the gastrovascular cavity of the stem, and many of them were released shortly after the specimens had been collected.
The Doc Ricketts Dive 500 specimen contained numerous stem fragments featuring several palpons, all typically with an oil droplet apposed to the proboscis. Their attachment was usually laterally, but occasionally they appeared to be at a 45° angle such that each proboscis projected outwards at a similar angle. There was no evidence to suggest that there was more than one palpon per cormidium.
Gonophores: ( Figures 38–40 View FIGURE 38 View FIGURE 39 View FIGURE 40 ). Both male and female gonophores were present on each specimen, although the holotype possessed very few of them. They were more numerous on the JSL 1405 and Doc Ricketts Dive 500 specimens. The male ones ( Figure 40 View FIGURE 40 ) measured up to 1.2 mm in length, and 0.7 mm in diameter and had a distinct spadix, and were generally smooth-walled. The female ones were smaller and spherical, measuring c. 0.25 mm in diameter, and the external ectoderm often showed the presence of large spherical cells. The pieces of siphosome were too contracted to be able to discern whether each cormidium possessed gonophores of both sexes, or were unisexual.
Remarks. As mentioned above Cordagalma bimaculatum sp. nov. is one of three Cordagalma species that are now known to occur in the western Mediterranean. The best known species, C. ordinatum is easily distinguished from the others by the presence of a rostrum on the larval-type tentilla. The differences between C. bimaculatum sp.nov. and the other Mediterranean species, C. isocarrei sp. nov, will be discussed below.
Distribution. The species is known from three widely separated sites; the Alborán Sea (western Mediterranean), the Bahamas, and Monterey Bay, California. In the warmer waters of the Mediterranean and the Bahamas the two specimens were found at similar depths, namely 747 and 726m, respectively. It should be noted that the specimen from the Mediterranean was inadvertently referred to as Cordagalma ordinata by Mills et al. (1996). In the colder waters of Monterey Bay the five specimens were found at much shallower depths, mainly between 400 and 587 m, with one specimen collected at 302 m. There are frame grabs for other in situ specimens of Cordagalma that were not collected but according to their depth probably were C. bimaculatum sp. nov. These were Tiburon Dives 1107 (404 m) and 1113 (470 m) and Doc Ricketts Dives 33 (494 m) and 104 (494 m).
Mackie (1985) made several observations of what he called Cordagalma cordiforme from British Columbian waters using the submersible Pisces IV. The bulk of these observations were within the 200–400 m depth range, with a few deeper records, but none shallower than c. 180 m. It is possible that some of the deeper observations refer to C. bimaculatum but, as we shall see, there are other Cordagalma species in this depth range that appear to be more common in Monterey Bay. Although Mapstone (2009) did not have any specimens of a Cordagalma species in her collections, she decided to include a description of C. ordinata = C. cordiforme = C. ordinatum on the basis that Mackie (1985) had noted its presence in British Columbian waters. The specimens used for this description were a couple belonging to the present author, of which one was actually C. bimaculatum sp. nov. Thus her figure 22D definitely represents a bract of C. bimaculatum sp. nov., as may those of the nectophores. The other bract was possibly that of C. ordinatum , although her interpretation of the nematocysts as "tubercles" is clearly incorrect.
Etymology. The specific name bimaculatum refers to the two patches of cells present on both the nectophores and the bracts.







| JSL |
JSL |
| MBARI |
MBARI |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Physonectae |
|
Family |
|
|
Genus |
Cordagalma bimaculatum
| P. R. Pugh 2016 |
Cordagalma ordinata
| Mapstone 2009 |
Cordagalma cordiforme
| Mills et al. 1996 |
Cordagalma cordiforme
| Mackie 1985 |