
Cordagalma ordinatum (Haeckel, 1888)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4095.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:690FFEBE-F71B-4EFD-865A-944D81A12897 |
|
DOI |
https://doi.org/10.5281/zenodo.6087811 |
|
persistent identifier |
https://treatment.plazi.org/id/03BC87D0-FF9F-FFD0-FF6E-F9C9FDA9D77A |
|
treatment provided by |
Plazi |
|
scientific name |
Cordagalma ordinatum (Haeckel, 1888) |
| status |
|
Cordagalma ordinatum (Haeckel, 1888) .
Synonymies: For more recent papers only those that describe the species in some way have been included.
Anthemodes ordinata Haeckel, 1888 a, p. 40; 1888b, p. 229, Pls. XIV, XV; Bedot, 1896, p. 409; Chun, 1897, p. 104; Schneider, 1898, p.117; Delage & Herouard, 1901, p. 225, fig. 381; Stepanjants, 1967, p. 89.
Cordagalma cordiformis Totton, 1932 , p. 325, figs. 8–9; 1954, p. 69; 1965, p. 61, fig. 25; Totton & Fraser, 1955, p. 3, fig. 10; Carré, C., 1968, p. 79, Pls. 1–3 [in partim] [non Pl. II, figs. 1?, 2,?3; P. III, figs. 1, 4, 7]; Carré, D., 1973, p. 113, fig. 1, Pls. 1–2; Palma, 1973, p. 27, Pl. II; Daniel, 1974, p. 49, fig. 3, K–N; 1985, p. 77, fig. 17; Stepanjants, 1977, p. 60, pl. 3a,b; Carré & Carré, 1980; p. 115, fig. 13; 1995, p. 565, figs. 171C, 175B, 181A, 188, 190; Casanova, 1980, p.21; Purcell, 1984, p. 319, fig. 4g, h, I; Mackie, 1985, p. 753; Gili, 1986, p. 268, fig. 4.49b, 4.63j,o; Mackie, Pugh & Purcell 1987, fig. 47; Pagès, 1991, p. 311, fig. 8.8; Pagès & Gili, 1992, p. 72, fig. 7;? Margulis, 1993, p. 17, Pl. 2a–g; Kitamura, 1997, p. 20, fig. 11; Pugh, 1999, p. 481, figs. 3.9, 3.24; Zhang, 2005, p. 22, fig. 7; Bouillon et al, 2006, p. 443, fig. 208A–B; Pugh, 2006b, p. 33.
Cordagalma cordiforme Cairns et al., 1991 , p. 33; 2002, p. 29; Pugh, 1999, p. 481, fig. 3.9 [non fig, 3.24]; Dunn, 2005, p. 33; Dunn et al, 2005, p. 924, figs. 5–8; Cartwright et al., 2008, p. 4; Cartwright & Nawrocki, 2010, p. 463;
Cordagalma ordinata Mills et al., 1996 , p. 154; Bouillon et al., 2004, p. 211, fig. 211A,B; Pugh, 2006b, p. 33;
non Cordagalma cordiformis Mackie, 1985 , p. 753.
non Cordagalma ordinatum Mapstone, 2009 , p. 109, fig. 22a,b,d.
? Cordagalma cordiformis Araujo, 2006 , p. 76, pl. 8, fig. 6 (non fig, 6.3).
? Cordagalma ordinata Araujo, 2012 , p. 69.
Diagnosis. Small nectophores with relatively short and narrow basal process. Shallow apical indentation between axial wings; nectosac, in ostial view square, with rounded corners. No distinctive patches of large, darkly staining, ectodermal cells on the main body of the nectophore, but small patches, usually 3 pairs, on the lower lateral sides of the velum of the ostium. Bracts rhomboidal with weakly defined truncated upper surface. Transverse and median ridges bearing nematocysts. Bracteal canal extending onto upper side of bract proximally, and then running along the lower side to the mid-length of the bract, ending in a small swelling inflected into the mesogloea. Palpacleless palpons attached by their bases. Tentilla with terminal rostrum.
Material examined. All specimens were collected by SCUBA divers at superficial depths. BWP stands for "Blue Water Plankton" and the following numbers represent a unique dive number, according to a system used during early diving cruises out of WHOI. "OC" stands for the WHOI research vessel Oceanus together with its cruise number.
BWP447 No station data available
BWP544-8 No station data available
BWP 567-15 No station data available
BWP 752-14 No station data available
BWP 781- 18 21-Jun-1979 31° 45'N, 71° 49'W GoogleMaps BWP 812- 13 13-Jul-1979 38° 57'N, 64° 35'W GoogleMaps BWP 816- 6 15-Jul-1979 40° 14'N, 68° 07'W GoogleMaps BWP 1070-7 26- Jul-1983 08° 37.3'N, 40° 22.4'W GoogleMaps BWP 1886-19 No station data available
OC368- 22-16 12-Jul- 2001 37°59'N, 72°58.9'W GoogleMaps OC368- 25-1 13-Jul-2001 37° 57'N, 73° 56'W GoogleMaps OC368- 25-8 13-Jul-2001 37° 57'N, 73° 56'W GoogleMaps OC368-25-19A 13-Jul-2001 37° 57'N, 73° 56'W GoogleMaps OC368- 26-9 13-Jul-2001 37° 54'N, 73° 00'W GoogleMaps OC368-32- 13 15-Jul-2001 39° 33'N, 71° 30'W GoogleMaps OC370- 6-6 20-Sep-2001 38° 41.83'N, 72° 54.48'W GoogleMaps
In addition, a poorly preserved specimen collected at Villefranche-sur-Mer by Mr A.K. Totton on an unspecified date was briefly examined.
Description. Pneumatophore: The minute pneumatophore ( Figure 4 View FIGURE 4 ) measured c. 0.3 mm in length and 0.12 mm in diameter. There was no notable pigmentation on the preserved specimens, but Carré (1968) said it was lightly pigmented apically, although no colour was mentioned.
Nectosome. The nectophores were budded off on the ventral side of the nectosome and the maximum number found with any of the preserved specimens was 16. Carré (1968), however, noted the presence of up to 40 nectophores on the nectosome of some of his specimens, but we cannot be sure that he was referring to this species (see Remarks section below).
Nectophore: The preserved nectophores measured up to 3.5 mm in length and 3.25 mm in width. However, although individual specimens tended to have fairly uniformly sized nectophores their mean size varied. Thus, the Oceanus 368 Dive 25- 1 specimen had smaller nectophores, averaging 2.32 mm in length and 1.92 mm in width; while the BWP 544- 8 specimen had larger nectophores averaging 3.37 mm in length and 3.02 mm in width. Another feature was that the younger nectophores tended to be broader than long, but as they grew they increased more in length than in width. Carré (1968) stated that his nectophores measured up to 7 mm in length and 6 mm in width; the implications of which again are discussed in the Remarks section below.
The hydroecium was wide on the main body of nectophore, but narrowed toward the basal process and petered out about halfway down the latter. No patches of distinctive large ectodermal cells or any sign of pigmentation were seen on the main body of the nectophore. In his notes, Claude Carré recorded the presence of spots of pigmentation on the nectophores, although he made no mention of them in his publication. Where these spots were located was also not made clear.
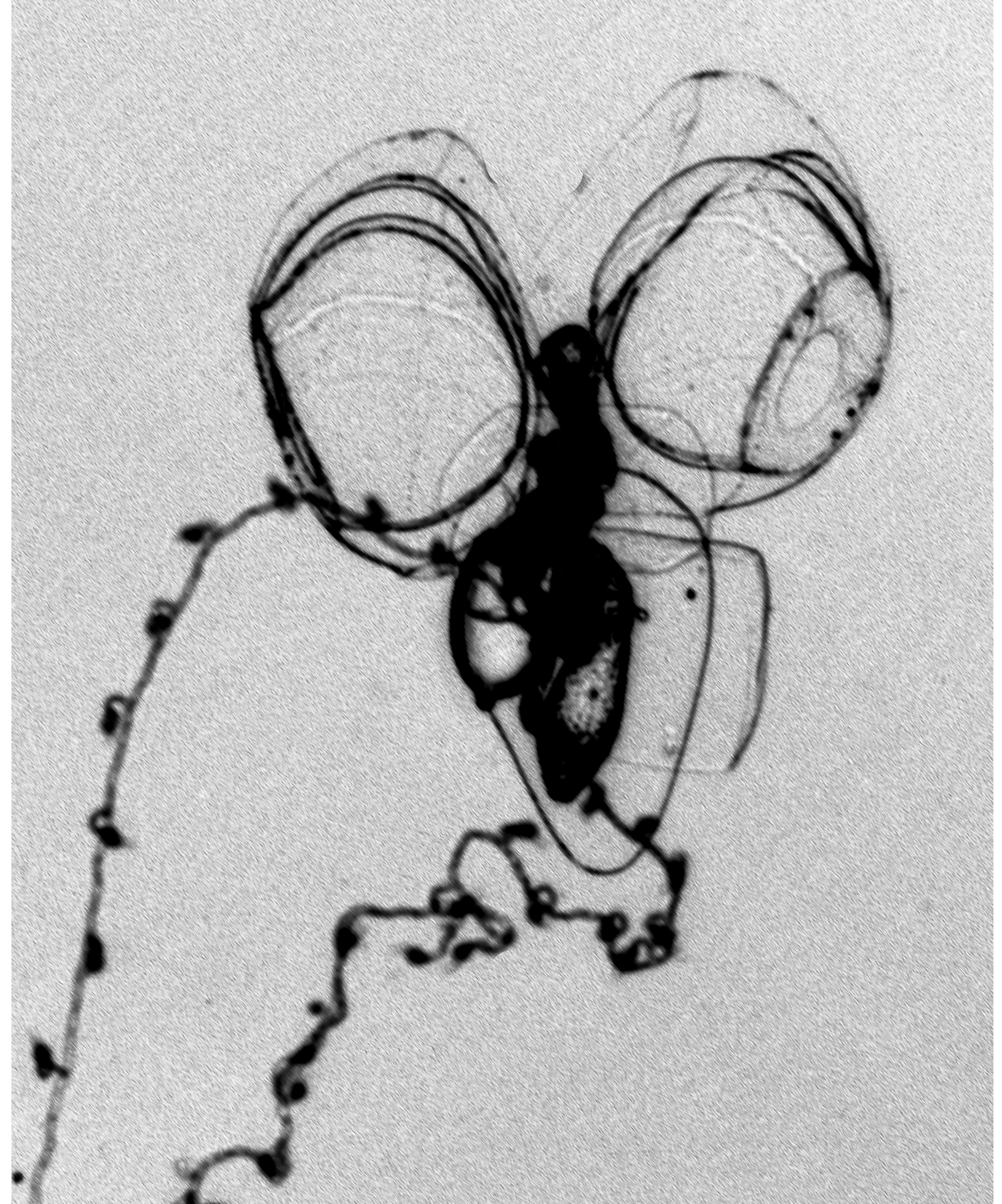
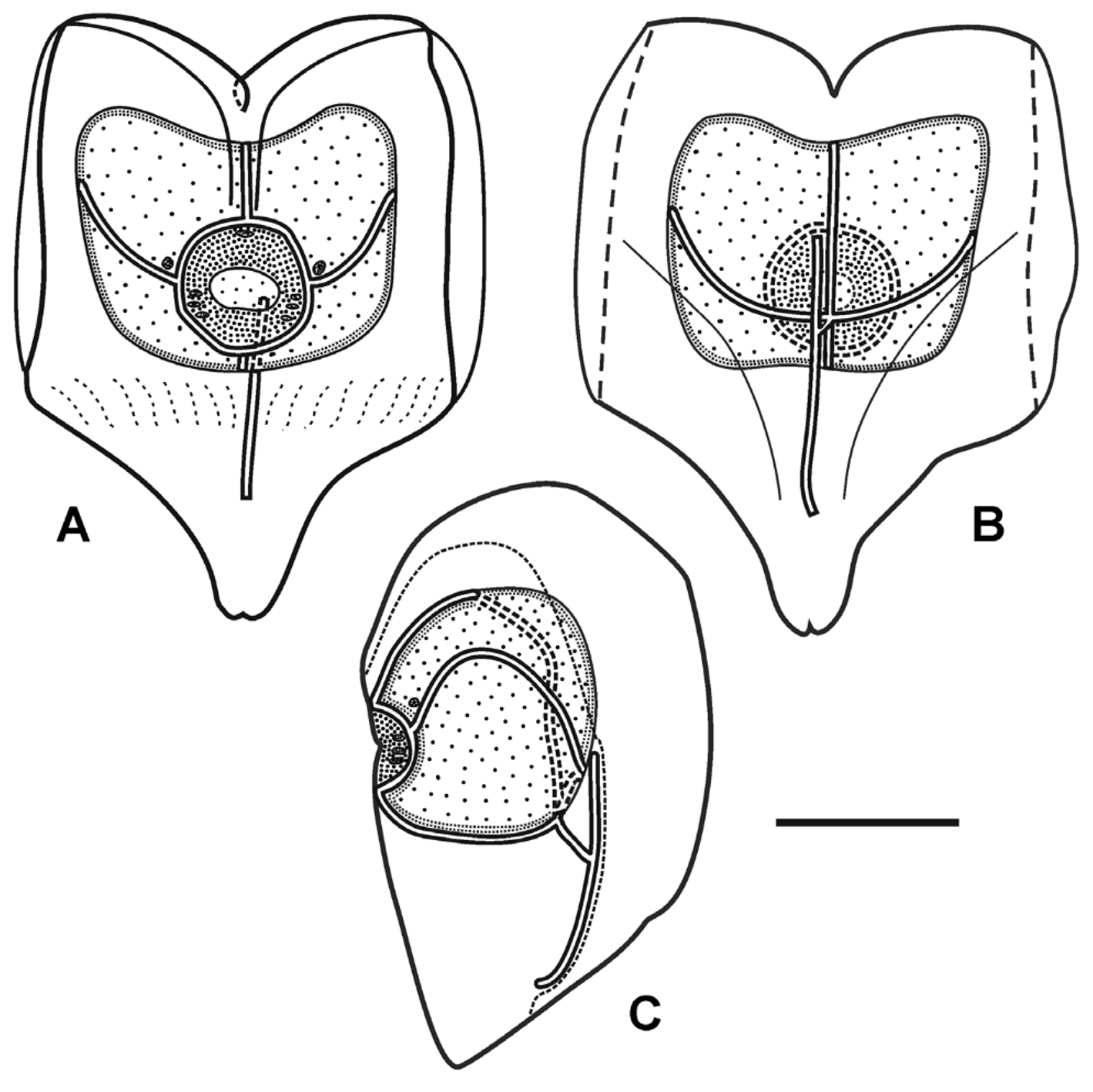
The nectosac was relatively small and did not extend basally, in the mid-line; with broad, shallow apical median indentation. Long ascending and descending mantle canals of approximately equal length. On the nectosac, the pedicular canal gave rise to all four radial canals. The dorsal and ventral canals ran straight to the ostial ring canal. The lateral canals, as is characteristic of cordagalmatid species, looped up and over onto the sides of the nectosac, then looped downwards, before running to the ostial ring canal ( Figure 5 View FIGURE 5 ). Typically their were three pairs of patches, made up of a few large ectodermal cells, on either side of the ostial velum at the level of, and below, the points of insertion of the lateral radial canals onto the ostial ring canal ( Figure 5 View FIGURE 5 A,C). Occasionally there were four pairs of patches, but often they had been abraded. Also, a patch of cells, sometimes divided into two or more, was frequently found on the velum below the point of insertion of the upper radial canal ( Figure 5 View FIGURE 5 A). Whether these patches of cells, which are probably sites of bioluminescence, also were pigmented in life is not recorded. In addition, there was a small, discrete patch of small cells lying over the end of each lateral radial canal but, unlike the lateral ostial processes found on the nectophores of other physonects these patches did not connect to the ostium itself.
The young nectophore ( Figure 6 View FIGURE 6 ) had a short rounded basal process. The axial wings were very small and only a very shallow emargination separated them. There was a pair of marked ridges that formed the outer margins of the overhanging hydroecial flaps. They ran from the outer margins of the axial wings towards the mid-line and then down to the level of the distal end of the descending mantle canal. In that region there was a broad, but shallow trough between them. Slightly lateral to these ridges there was a row, or two, of nematocysts that ran from the axial wings to about half the height of the nectophore. Further rows of nematocysts were found on the ostial side of the nectophore, but these were not associated with distinct ridges. No lateral patches of cells could be discerned on the velum of the nectosac, but there usually was one on its upper side.
Siphosome: Haeckel (1888b) and Carré (1968) both noted that several hundred cormidia could be found on a single specimen, and that each cormidium consisted of, from posterior to anterior, a gastrozooid, a palpon without palpacle, a bunch of male, a bunch of female gonophores, and 6–8 bracts, which budded from the bases of the other zooids. However, the latter author also found that for some of his specimens the gonophores of individual cormidia were of only one sex, although both sexes were present on the whole colony. Presently it is not possible to clarify without doubt whether this is a specific character or, perhaps, whether it suggests that the specimens are protandrous or protogynous.
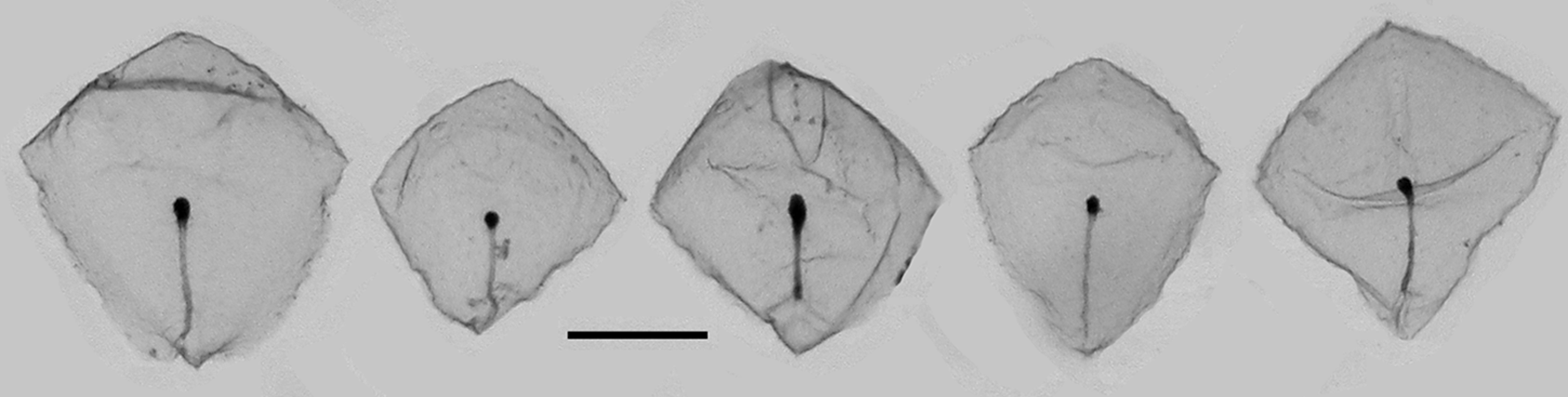
Bracts: For the present specimens the preserved bracts ( Figures 7–9 View FIGURE 7 View FIGURE 8 View FIGURE 9 ) measured up to c. 1.25 mm in length and 1.125 mm in width. They were roughly rhomboidal in shape, but the truncated part of the upper surface was only weakly defined. There was a pronounced transverse ridge and a median one that divided the distal facet into two. No patches of ectodermal cells were found on these facets. The canal extended onto the upper side of the bract and curved over its proximal end to run, in the mid-line, along its lower side. It ran to about the mid-length of the bract where it ended in a small swelling inflected into the mesogloea. This inflection was more pronounced in the younger bracts ( Figure 8 View FIGURE 8 ).
Carré (1968, p. 81) described the bracts as having "la form d'une pyramide tronquée assez régulère, avec quatre faces latérales et des crêtes garnies de petit nématocytes" [the form of a quite regularly truncated pyramid with four lateral facets and its ridges garnished with nematocysts]. The nematocysts were scattered along the transverse and median ridges ( Figures 5–6 View FIGURE 5 View FIGURE 6 ) but more often than not they had been abraded. They were not investigated in detail for the present study, but Carré (1968) identified those on the "distal ridge" as being small stenoteles measuring 8.5 x 7.5 µm, while on the other ridges they were probably atrichous isorhizas. Although Carré's description of the bracts clearly indicates them as belonging to Cordagalma ordinatum , care must be taken when identifying the bracts from his photographs as those shown in his Plate II, fig. 2 and Plate III, figure 1 certainly do not belong to that species, although those in Plate III, fig. 5 might. (For further discussion see Remarks section).
Larval bract: No larval bracts were found with the present specimens examined, but Carré (1973), who followed the development of Cordagalma ordinatum , showed that a simple, almost circular caducous larval bract was developed before the nectophores appeared, but she gave no details about it, including its size. It appears that it is retained until at least the first pair of nectophores has been developed (see Figure 4 View FIGURE 4 ).
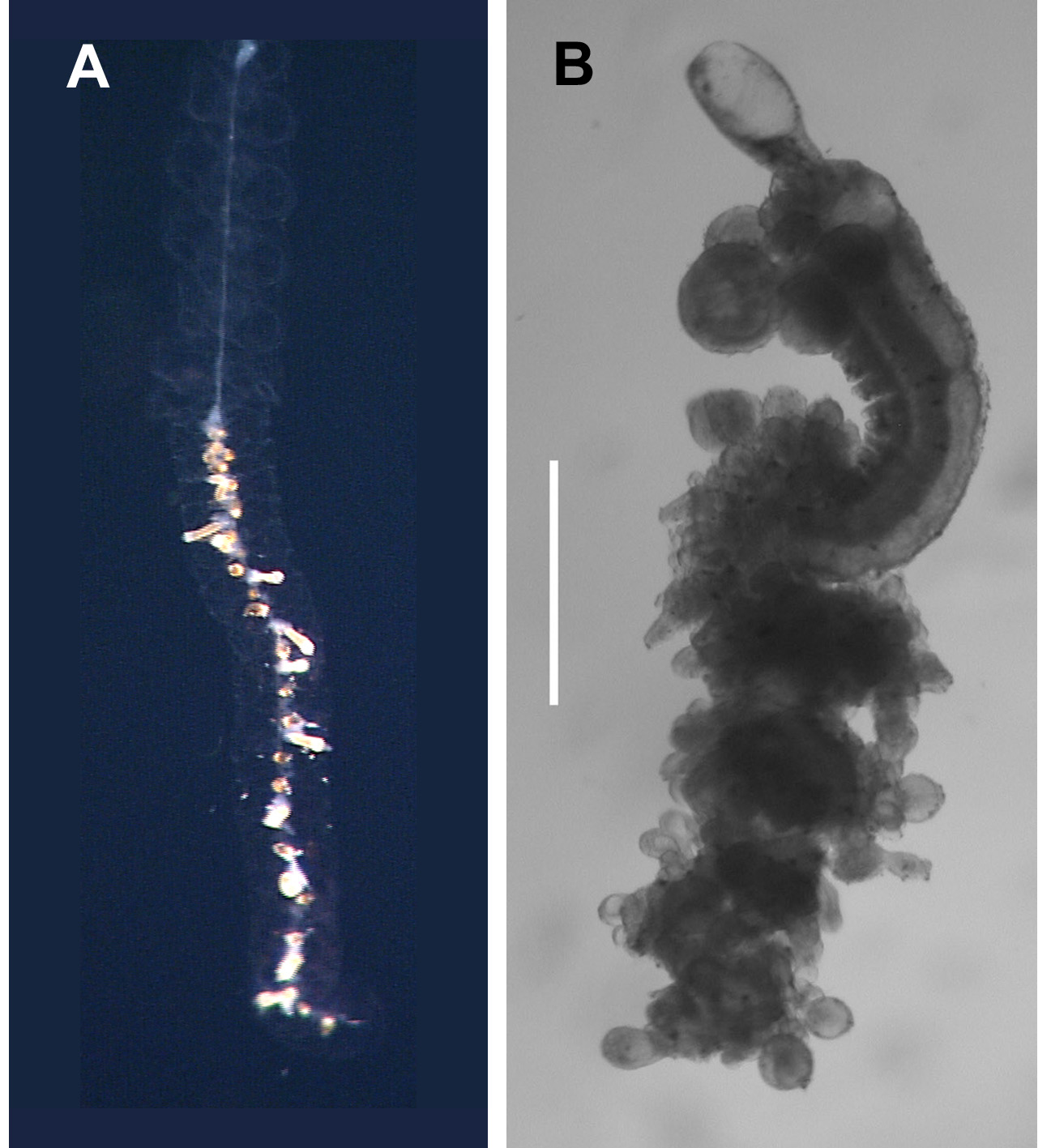
Gastrozooid: The siphosome of the preserved specimens frequently broke into fragments consisting of a gastrozooid, a palpon, and one or two gonodendra ( Figure 10 View FIGURE 10 ), with or without bracts. The unpreserved gastrozooids ( Figure 10 View FIGURE 10 D, gz) typically measured c. 2 mm in length and were connected to stem by a short peduncle. They were transparent and colourless except, according to Carré (1968), for two orange-red spots at their bases but these were not apparent on the current photographs. The distal proboscis section was quite long as shown by the length of the narrowed gastrovascular cavity. Hepatic stripes were present in the slightly expanded stomach region; while the basigaster was very small. After preservation ( Figure 10 View FIGURE 10 A–C) the gastrozooids became opaque. They usually featured a pronounced proboscis region, occupying approximately a third of the total length, a slightly expanded stomach that was separated from the now very distinctive basigaster by a constriction, from one side of which the tentacle arose.
Tentilla: The tentacles could extend to considerable lengths and Carré (1968) recorded up to 100 tentilla on each, but such was impossible to gauge from the present fragmented preserved material. Each tentillum, when uncontracted, had a thin pedicle, but this was always highly contracted in the preserved ones ( Figure 11 View FIGURE 11 A). The cnidoband formed an ovoid structure that contained three types of nematocyst. Proximally there was an incomplete ring of five large stenoteles that were somewhat variable in size. Carré's measured 15.5 x 8.5 µm, while larger ones, which measured 20 µm in length and 10 µm in diameter, were found with the present material. In the central part of the tentillum there were numerous anisorhizas (16 x 2 µm), and a few isorhizas, possibly holotrichous, measuring 5.5. x 5 µm.
Strangely, Carré (1968) did not describe the distal part of the tentillum that consisted of several very long cnidocils stretching out in 2π directions, nor the character that, as will be shown, sets Cordagalma ordinatum apart from all other cordagalmatids, the "rostre" or rostrum. In our preserved specimens the "rostre" tended to be contracted and difficult to see ( Figure 11 View FIGURE 11 A), although occasionally it was easily distinguished ( Figure 11 View FIGURE 11 B). No nematocysts were present on the rostrum. The cnidocils also appeared much shrunken and distorted ( Figure 11 View FIGURE 11 A). These cnidocils are presumed to be part of the isorhizas of the cnidoband, although this has not been confirmed. Carré's photograph (1968, Plate III, figure 6) clearly shows all the features mentioned above but, unfortunately, it was not one of the ones he sent to me.
Palpon: The fully developed palpons were c. 1 mm in length ( Figure 10 View FIGURE 10 ). In life they consisted of a long pedicle (see Carré, 1968, Plate II, figure 4; but not Plate III, figure 4), a globular stomach region and a short narrow distal proboscis ( Figure 10 View FIGURE 10 D). The pedicle was inserted into the base of the stomach region so that its attachment was proximal. This arrangement could remain unaltered on preservation ( Figure 10 View FIGURE 10 C), but more often than not considerable changes in form occurred. The stomach region becomes more elongate and cylindrical but, in particular, the pedicle contracts in such a way that it appears to be attached to one side at the base of the stomach region ( Figure 10 View FIGURE 10 A, B). This, possibly, refers to the intermediate attachment position that Carré referred to (see below). There was never any sign of the attachment of a palpacle.
Gonophores: The gonophores were extremely small and, although it was possible to distinguished the male ones (see Figure 10 View FIGURE 10 ), it was extremely difficult positively to identify the female ones. It is thought, by its structure, that the one shown in Figure 10 View FIGURE 10 B is female, but only based on the very broad anastomosing radial canals that were present. No gonophores of the sort illustrated by Carré (1968, Plate III, figs. 2 & 3) were ever observed, nor did Carré mention the presence of anastomosing radial canals.
Remarks. Although, as noted above, the nectophores of Anthemodes ordinata described and illustrated by Haeckel (1888b) bear no resemblance to those of Cordagalma ordinatum , once a whole specimen had been, albeit briefly, described by Carré (1968), it was possible to note that their descriptions of the siphosomal elements bore close similarities.
It is not necessary to detail exhaustively these similarities here but both Haeckel and Carré noted the simplicity of each cormidium, with its gastrozooid, with a tentacle, a palpon, cyston or dactylozooid without palpacle, a male and a female gonodendron and a number of bracts. Such similarities can also be found in their descriptions of the bracts, and the presence of nematocysts on the margins of the facets; the types and distribution of nematocyst on the tentillum, and the presence of a rostrum, belatedly mentioned by Carré, and mistakenly referred to by Haeckel as a terminal filament despite the fact it bore no nematocysts.
Despite the brevity of Carré's (1968) description there are six brief paragraphs, under the heading "Variabilité" that are of great importance. Apart from some variations in pigmentation that may only be resolved by examining living material, the most important variations in characters were:
a) In some specimens the pedicle of the palpon was attached laterally, instead of the more usual basal positioning, with some being intermediate between the two;
b) in some specimens a terminal "rostrum" on the tentillum was absent; and
c) in some specimens the individual cormidia included gonophores of only one sex, whereas in others gonophores of both sexes were included.
Carré (1968) considered that it would be premature to consider that these novel characters were of sufficient importance to warrant the designation of a new species mainly, it appears from his notes, because he was unable to be sure that the three characters together formed two distinct groupings.
Having looked at more recently collected specimens of Cordagalma ordinatum , I conclude that the first two of these characters do, in fact, form two distinct groups, and that they provide sufficient evidence to divide Carré's specimens into two different species. C. ordinatum has palpons that are attached by their bases to the stem, although there are some differences between the living and preserved material (see above). In addition their tentilla have a terminal "rostre". Thus, it seems that the lateral attachment of the palpons and the absence of a "rostre" on the tentillum are sufficient to distinguish a different species. Fortunately, specimens with very much the same characters, allowing for differences between Carré's fresh and our preserved material, have been collected in Monterey Bay, California and are described herein under the name C. isocarrei sp. nov. Comparisons will be made between that material and some unpublished pictures to which Claude Carré has kindly given me access.
Thus, one must be very careful when using any of Carré's (1968) description, and particularly his photographs, for identifying Cordagalma ordinatum . For instance, although the figures on Plate I probably all refer to C. ordinatum , at least two, and probably three, of those on Plate II refer to the new species.
Distribution. While Haeckel's (1888b) original specimen came from the Canary Islands that of Totton (1932) came from the Great Barrier Reef. The species has clearly been found in the superficial waters of the Western Mediterranean (e.g. Carré, 1968; Dallot et al., 1988; Gili et al., 1987) while there are no records for any Cordagalma in the eastern Mediterranean. However, C. isocarrei sp. nov. also occurs in the western Mediterranean, probably at quite shallow depths, based on the records from Monterey Bay, and a third species has been found in the Alborán Sea, which Mills et al. (1996) mistakenly referred to as Cordagalma ordinata .
Cordagalma ordinatum has frequently been found in the Sargasso Sea (N.W. Atlantic Ocean) (personal information). However, with so many new species herein described, it is difficult to be certain that the relatively sparse number of records for a Cordagalma species definitely refer to C. ordinatum . It appears to be a species that lives in superficial warmer waters, at least in the Atlantic Ocean, although its presence in other Oceans remains to be confirmed, with certainty. The record of Pagès et al. (2006) from Oceanographer Canyon, off Woods Hole in the North Atlantic might be dubious, although it was collected at the surface, while records from warmer waters (e.g. Biggs, 1977; Purcell, 1981; Dunn, 2005), including the Gulf of Mexico (Burke, 1975; Gasca, 1993) are likely to be of C. ordinatum . The only deep record for a Cordagalma species comes from Casanova (1980) from the 500–700m depth range near Great Meteor Bank (c. 30°N, 30°W). This might indicate that there was either surface leakage into the net, or that the species was not C. ordinatum .
In the South Atlantic potentially Cordagalma ordinatum has only been recorded in the Benguela current system off South Africa ( Pagès & Gili, 1992). There are only occasional records for the species in the Indian Ocean and the Red Sea (e.g. Totton, 1954; Daniel, 1985), while in the south Pacific Ocean, it has been collected from the Great Barrier Reef, of Australia, by Totton (1932) and Russell & Colman (1935), in eastern and central regions ( Stepanjants, 1967; Margulis 1993), and in the eastern region, off Chile (e.g. Palma, 1973). In the northwest Pacific it has been collected in Japanese ( Kitamura et al., 2003) and Chinese ( Zhang, 2005) waters.
In the northeast Pacific the only records for Cordagalma ordinatum appear to come from the colder waters of British Columbia, Canada (e.g. Mackie, 1985). However, the extensive studies, using ROVs and SCUBA diving, by Dr. S.H.D. Haddock and colleagues in the Monterey Bay region, northern California, have not revealed the presence of C. ordinatum , although several other Cordagalma species are present there, as described herein. It is, therefore, suspected that the records from British Columbia refer to one of the new species described below. Certainly Mapstone (2009, Fig. 14 View FIGURE 14 B) does not belong to C. ordinatum as the tentillum has no rostrum; the structure referred to as a rostrum actually represents the distal nematocysts, with their long cnidocils. Similarly, the nectophore and bract shown in her Fig. 22A View FIGURE 22. A , B, D actually belong to one of the new Cordagalma species described herein, as they were apparently drawn from one of my specimens of that species.
There are also a few other records for Cordagalma ordinatum that probably refer to new Cordagalma species. For instance, Pagès et al. (1996) recorded the presence of some specimens of C. cordiformis in Hardangerfjord, Norway and, similarly. Hosia & Båmstedt (2007) collected 27 specimens of what they called C. ordinata in the Korsfjord, Norway, using nets fished from close to the bottom (c. 600m) to the surface; and a single specimen in Fanafjord (0–150m). Since no specimens of C. ordinatum appear to have been collected in the northeast Atlantic Ocean at latitudes greater than that of the Straits of Gibraltar, it is felt that the Norwegian specimens most probably belong to a new Cordagalma species. Unfortunately, like those of Sphaeronectes sp. (see Pugh, 2009), these specimens were not retained by the authors and so their identification cannot be confirmed.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Physonectae |
|
Family |
|
|
Genus |
Cordagalma ordinatum (Haeckel, 1888)
| P. R. Pugh 2016 |
Cordagalma ordinata
| Araujo 2012 |
Cordagalma ordinatum
| Mapstone 2009 |
Cordagalma cordiformis
| Araujo 2006 |
Cordagalma ordinata
| Mills et al. 1996 |
Cordagalma cordiforme
| Cairns et al. 1991 |
Cordagalma cordiformis
| Mackie 1985 |
Cordagalma cordiformis
| Totton 1932 |
Anthemodes ordinata
| Haeckel 1888 |