
Cordagalma isocarrei, P. R. Pugh, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4095.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:690FFEBE-F71B-4EFD-865A-944D81A12897 |
|
DOI |
https://doi.org/10.5281/zenodo.6087823 |
|
persistent identifier |
https://treatment.plazi.org/id/03BC87D0-FFBC-FFEF-FF6E-FB63F8CED2D5 |
|
treatment provided by |
Plazi |
|
scientific name |
Cordagalma isocarrei |
| status |
sp. nov. |
Cordagalma isocarrei sp. nov.
Diagnosis. Small nectophores that, when mature, have a relatively short, broadly truncated basal process. A pair of small lateral ostial processes and, usually, a pair of patches of ectodermal cells on the lower lateral sides of the velum. No other patches present. Preserved specimens have a pair of furrows on the ostial side of the nectophores. The descending mantle canal is longer than the ascending one. Small rhomboidal bracts with more or less welldefined transverse and median ridges. The upper side is truncated; the distal margins formed by the transverse ridge, the proximal ones extremely vague. A pair of patches of ectodermal cells on the distal facets of younger bracts, but lost in the fully mature ones. Palpons are attached baso-laterally so that the proboscis and opening are at an angle of c. 135° to the stem.
Material examined.
Ventana 3557-D7 36°42'N 122°02'W 3-May-2010 Depth 214m GoogleMaps Ventana 3558-D2 36°42'N 122°02'W 3-May-2010 Depth 206m GoogleMaps Ventana 3558-D6 36°42'N 122°02'W 3-May-2010 Depth 218m GoogleMaps Ventana 3642-D1 36°42'N 122°02'W 11-May-2011 Depth 115m GoogleMaps Ventana 3629-D7 36°42'N 122°03'W 11-May-2011 Depth 357m GoogleMaps Ricketts 199-SS9 36°42'N 122°06'W 12-Oct-2010 Depth 421m GoogleMaps
Holotype: The specimen collected during the Ventana Dive 3558-D2 has been designated the holotype, and will be deposited at the United States National Museum (Smithsonian Institution), Washington, DC.
The remaining specimens will be placed in the collections of Dr Casey Dunn, at Brown University, Providence, Rhode Island, USA.
Description. A very poor in situ frame grab of the Ventana Dive 3558-D 6 specimen, but the best available, is shown in Figure 41 View FIGURE 41 . It is possible to see that the siphosome was very long in comparison with the nectosome (bottom right) and that the whole structure was very delicate.
Pneumatophore: The pneumatophore was minute, measuring c. 0.35 mm long and 0.14 mm in diameter. There were no obvious signs of pigmentation.
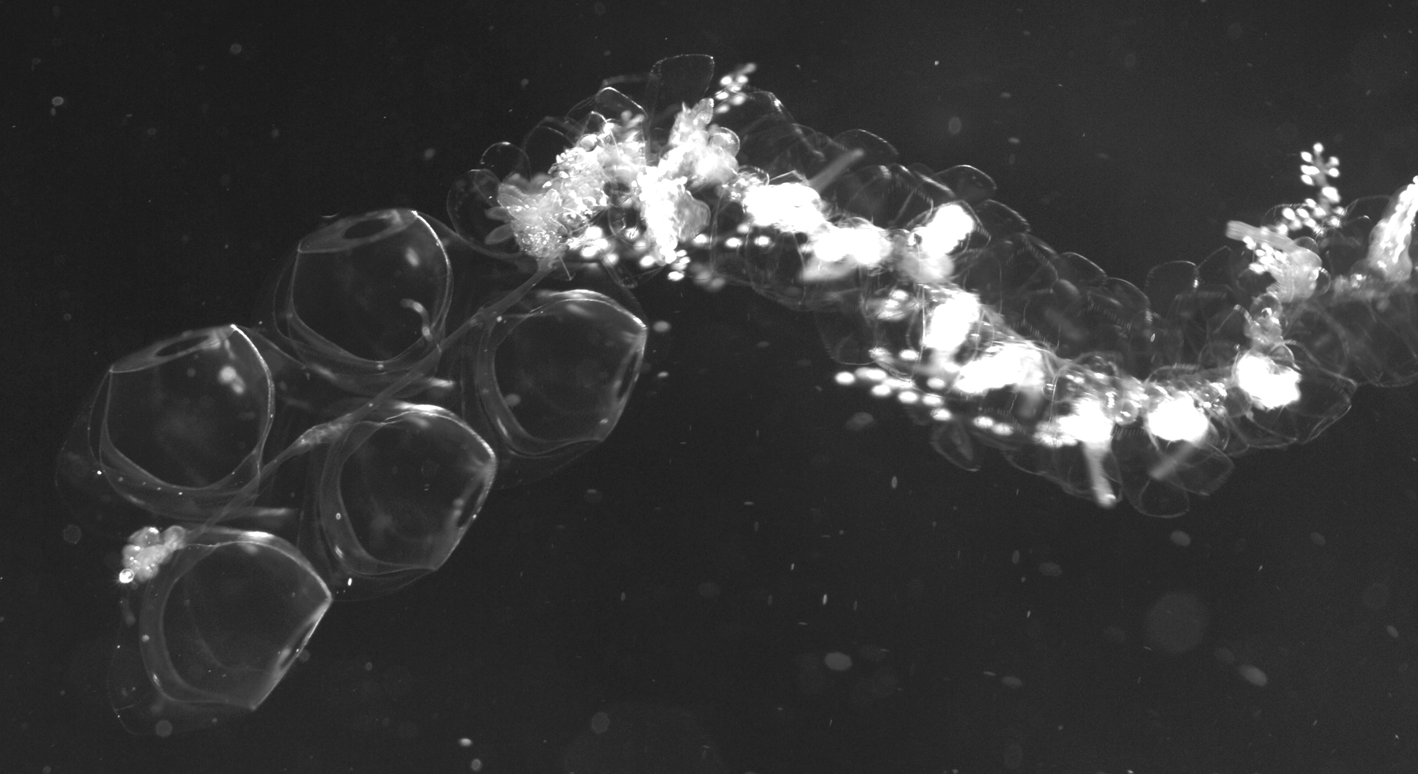
Nectosome: The nectosome was relatively short and the maximum number of nectophores retained with any specimens was fifteen mature, one developing and one very young. They were attached to the ventral side of the nectosome. Their alternating arrangement can clearly be seen in Figure 42 View FIGURE 42 , although the number of nectophores at that time had been reduced to five.
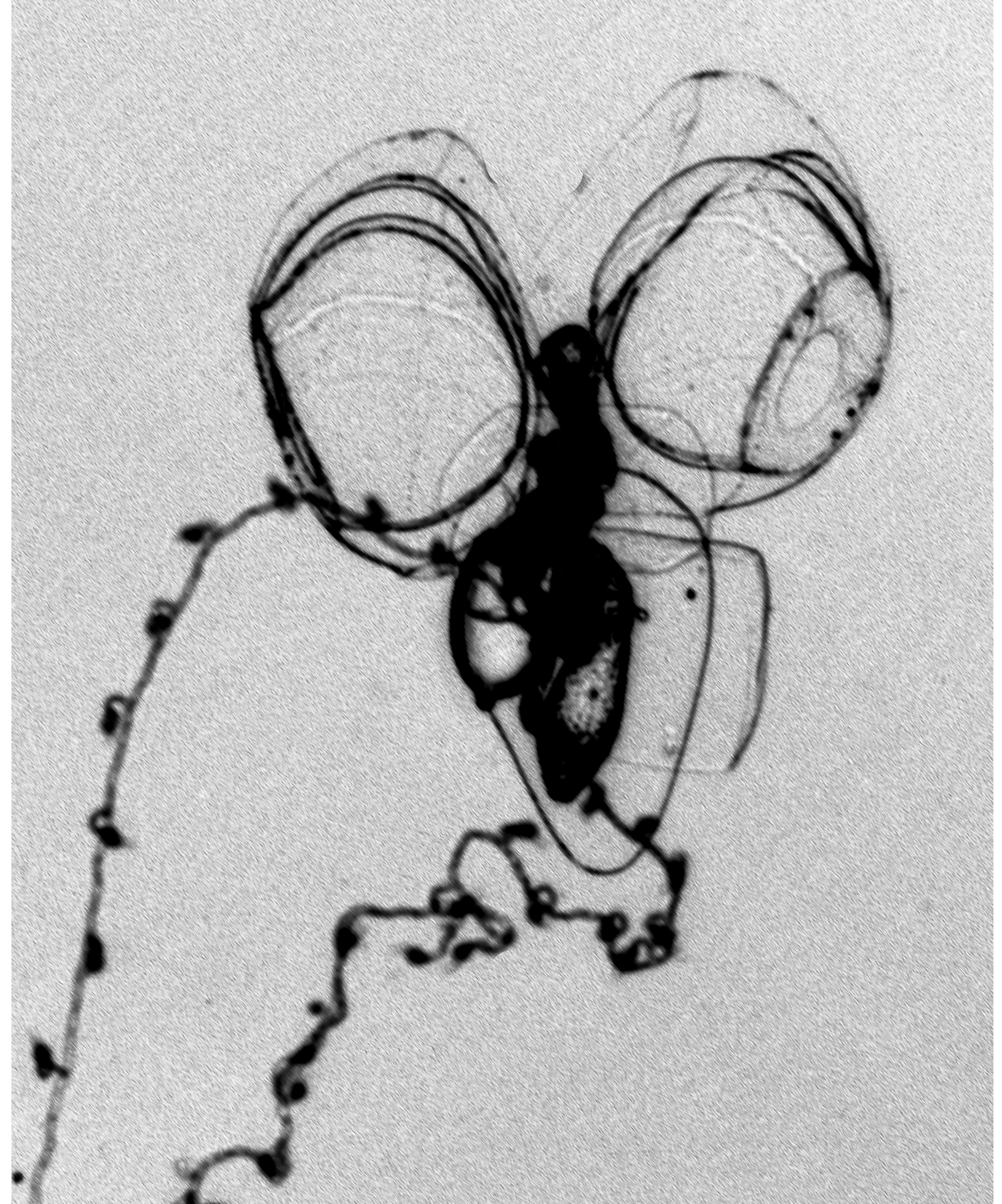
Nectophore: ( Figure 43 View FIGURE 43 ). Seventeen small nectophores were found with the type specimen, which was the largest number for any specimen. There was also a loose nectophoral bud. They measured up to 5 mm in length and 3.9 mm in width. On the older nectophores ( Figure 43 View FIGURE 43 A) the basal process was relatively short and broad, with a broad distal portion. However, in the developing nectophores ( Figure 43 View FIGURE 43 B, C) it was narrower and more pointed. The rounded axial wings were quite extensive and, usually, were separate by a narrow V-shaped indentation. On the ostial side of the nectophore, a pair of ill-defined ridges was present on the inner sides of the axial wings that continued down to above ostial level and demarcated a deep gutter between them. Also on the ostial side was a pair of lateral furrows that ran obliquely out and up from the lateral margins of the ostium. They may have been preservation artefacts, but their presence was quite constant.
On the axial side of the nectophores there was a pair of lateral ridges that demarcated the margins of the hydroecium. They were very vague in the upper half of the nectophore, but more prominent basally before they petered out just below the lower level of the nectosac. In the younger nectophores they tended to approach each other more closely, while in the larger ones the mesogloea on the inner sides was thickened with the narrow hydroecial furrow running between them.
The upper lateral margins extended into the axial wings to a greater or lesser extent, while the lower margin was almost squarely truncated. The radial canals had typical courses, the upper and lower being straight, the laterals first curving out and diagonally upwards, then down to join the ostial ring canal. The long descending mantle canal extended down into the basal process, although its distal portion was very vague. The ascending canal was shorter and extended up to the mid-ostial region. The pedicular canal gave rise to all four radial canals. Above each lateral radial canal, just before it joined the ring canal there was a small patch of ectoderm cells. Another pair of patches was present on the lower lateral sides of the velum. These patches were often extensive in the younger nectophores, but in the mature ones were smaller and more rounded.
Siphosome: Part of the long siphosome is shown in a tank photograph ( Figure 42 View FIGURE 42 ) of the V3629 specimen taken shortly after collection. For the preserved specimens the cormidia tending to separate into individual cormidia, sometimes more ( Figure 44 View FIGURE 44 ). These typically consisted of a posterior gastrozooid and tentacle, bearing tentilla, followed by a palpacleless palpon and gonophores of both sexes, although it is not clear in what order these were attached. Some photographs (e.g. Figure 44 View FIGURE 44 top left appear to show the male gonophores attached posterior to the female ones, while the reverse is possibly the case for others (e.g. Figure 44 View FIGURE 44 , bottom right).
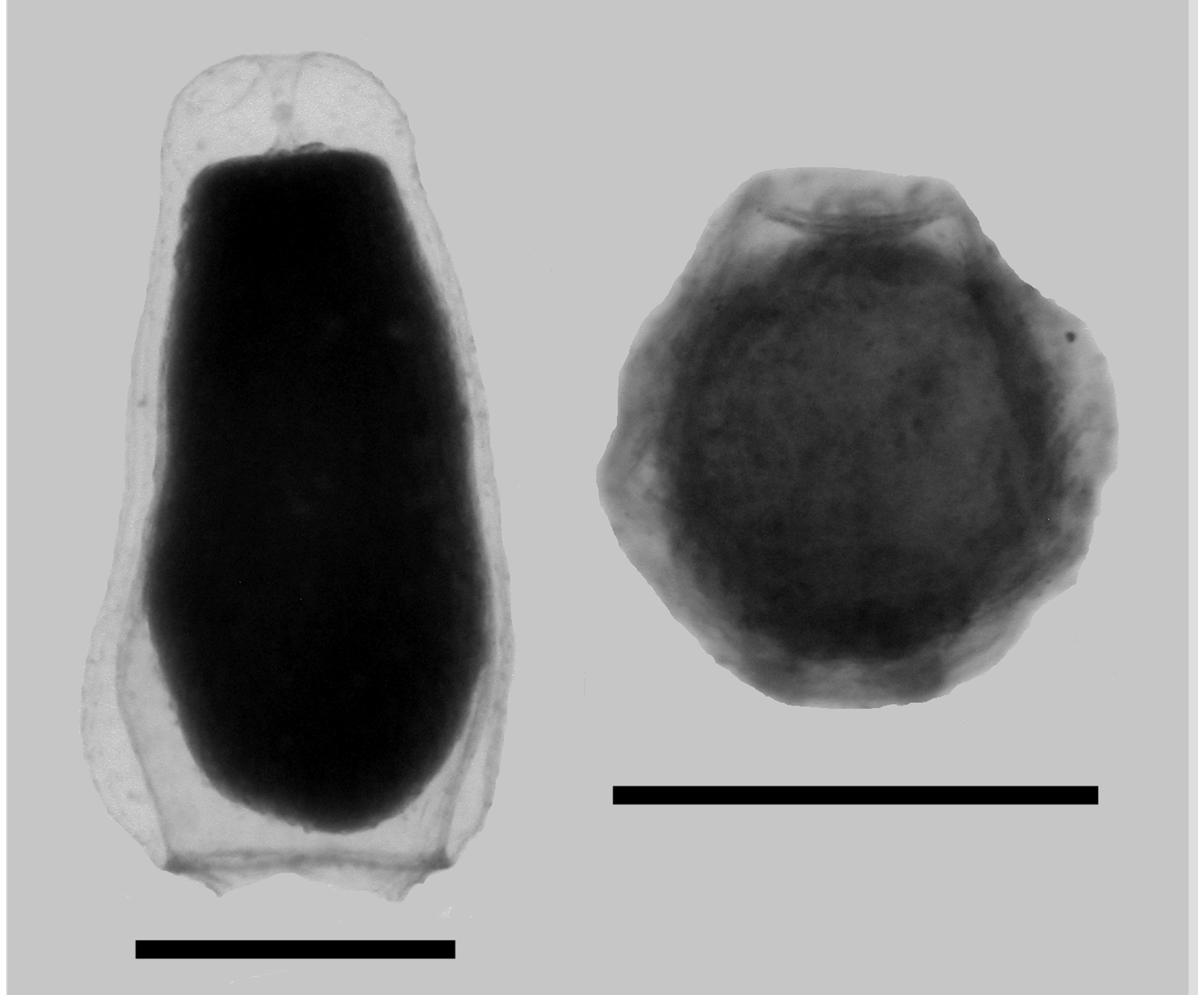
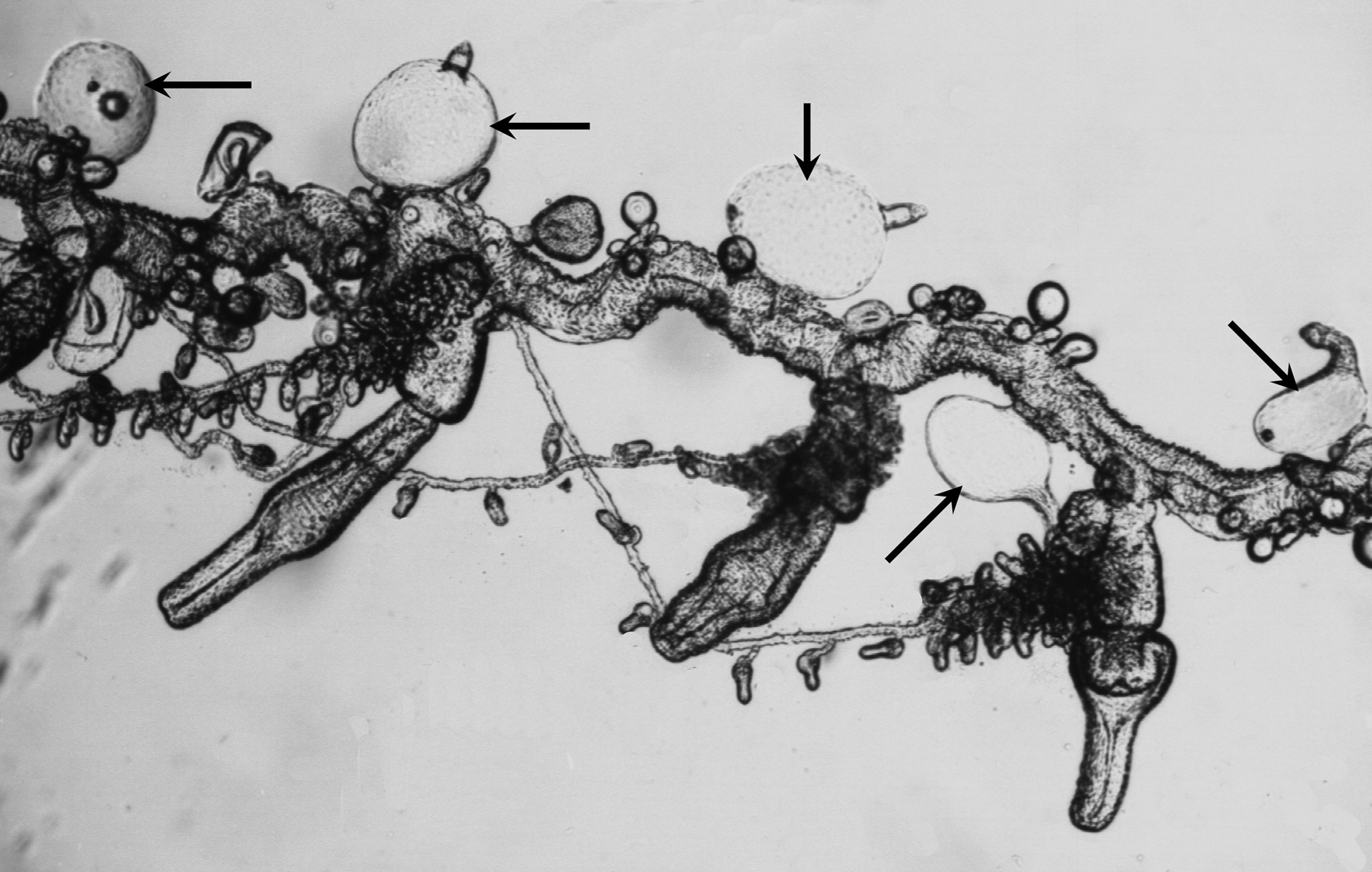
Bracts: The bracts ( Figure 45 View FIGURE 45 ) measured up to 2.2 mm in length and 1.8 mm in width. They were roughly rhomboidal in shape, with the younger ones resembling a diamond, the older ones more elongate. In the preserved state they were covered in pimples, but that was probably an artefact. The lower surface was flat, while the upper one was thickest centrally. There was a more or less well-defined transverse and median ridge; often with both appearing as just a rounded processes. Spherical nematocysts were found spaced out in a single row along the median ridge and, occasionally, onto the ridge demarcating the lower side of the distal facet. Also, on the younger bracts it was sometimes possible to see much smaller nematocysts arranged along the transverse ridge, but they had been lost in the mature ones.
The central part of the upper surface was truncated, although the proximal margins of this truncation were often extremely difficult to see. Occasionally it was found that the truncation narrowed and stretched down to the proximal end of the bract. The bracteal canal arose close to, but not at, the proximal end of the bract and stretched up to its mid-length. It was terminated by a large ovoid swelling that was inflected into the mesogloea. The connection between this and the canal itself was sometimes very difficult to see. On the younger bracts there was a pair of patches of ectodermal cells on the distal facets, either side of the median ridge. These gradually disappeared such that there was no trace of them on the mature bracts.
Gastrozooid: The gastrozooids ( Figure 44 View FIGURE 44 ) were small and quite variable in shape. They consisted of a long proboscis region, a more or less inflated stomach and a small, narrow basigaster. They appeared to be attached directly to the stem.
Tentilla: The mature tentillum ( Figure 46) measured c. 120 µm in length and 100 µm in diameter. It had the typical Cordagalma organisation with five large stenoteles proximally, which measured 47 x 22 µm, partially surrounding numerous anisorhizas and surmounted, distally by a few ovoid isorhizas, with extremely long cnidocils that often were difficult to discern because of their transparency.
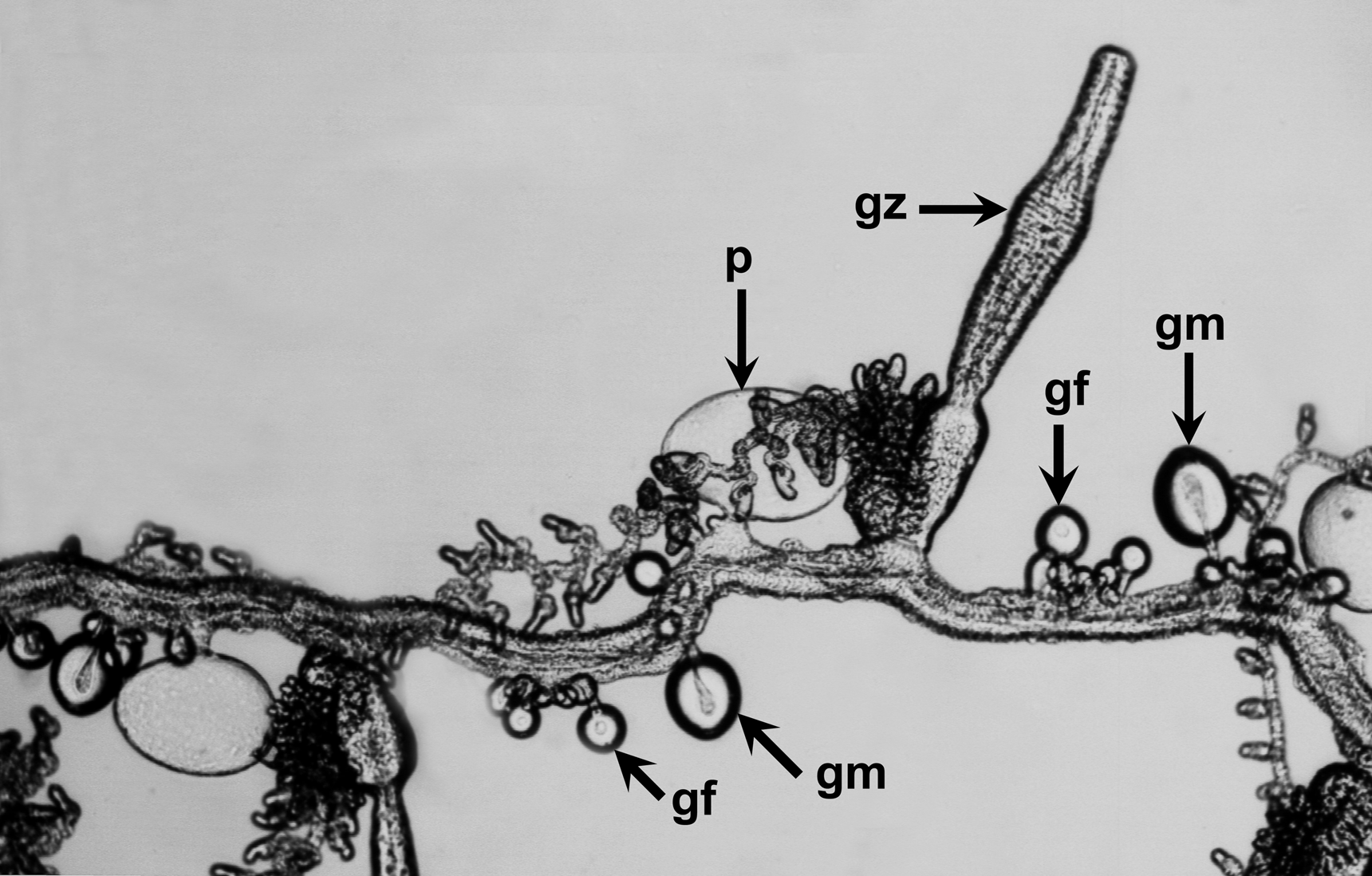
Developing tentilla had the typical form noted in other species and were not investigated further, but see below. Palpon: The palpons ( Figures 47–48 View FIGURE 47 View FIGURE 48 ) generally measured up to 1.7 mm in maximum width and 1.0 mm in height, and were attached to the stem by a very short pedicle. The attachment point was not at the proximal end of the palpon but slightly lateral to it, so that the proboscis region stuck out at an angle of c. 135° to the stem. The proximal end of the palpon was usually marked by the presence of an oil droplet, of variable size.
In a few cases ( Figure 48 View FIGURE 48 ) the palpons took on a very different shape. Although this might have been a preservation artefact, most of the palpons found with the Ventana Dive 3557 specimen were of this kind. The proboscis region became very pronounced in relation to the main body and they were filled with droplets of oil.
Gonophores: Mature gonophores of either sex were not often found with the specimens examined. However, the male ones ( Figure 49 View FIGURE 49 , left) were quite distinctive and measured c. 1.35 mm in length and 0.7 mm in maximum diameter. The female ones were always very small and it was not clear as to whether the largest, which measured c. 0.55 mm in length and diameter, were actually fully mature.
Remarks. Cordagalma isocarrei sp. nov. and C. bimaculatum sp. nov. are the only two species of Cordagalma where the palpon is known to be attached laterally and not proximally. However, the angle at which the proboscis region projects out from the main body of the palpon is quite different, being 135° in the former and 90° in the latter. The differences in the morphology of the bracts of these two species, as is discussed below, should, in association with the above, clearly distinguish the two. The mature tentilla of neither species possess a rostrum, and the other Mediterranean species, C. ordinatum , remains unique.
It appears that Cordagalma isocarrei sp. nov. is the solution to the enigma as to the identity of the second species that Carré (1968) apparently described. What we know for certain about that species is that the tentilla were without a "rostre" and that the palpons were attached laterally, although Carré (ibid p. 83) also noted "certaines colonies ont des dactylozoïdes de type intermédiaire à insertion baso-latérale" [certain colonies have an intermediate type of dactylozooid that is attached baso-laterally]. In addition, he noted that some of the cormidia included gonophores of only one sex but, in that case, we do not know to which species he was referring. As has been discussed in the description of C. ordinatum , it has been very difficult to establish whether the cormidia of that species were monoecious or dioecious. On the other hand, the two other characters clearly distinguish it from C. ordinatum .
Claude Carré took a number of photographs of his Cordagalma specimens and kindly provided some of these to me, together with some notes on his findings. Of particular interest are some fairly detailed observations and several photographs of a specimen that was collected on 14th March 1968. Having studied these in detail, we believe that there is sufficient evidence to consider that this particular specimen belongs to C. isocarrei sp. nov.
None of Carré's photographs of the nectosome and nectophores can be positively identified as either species, although from the pattern of patches on the velum it seems that that of a post-larval (see Figure 4 View FIGURE 4 ) probably belongs to Cordagalma ordinatum . His notes suggested that the two pigment spots present on the nectophores of C. ordinatum were absent in C. isocarrei sp. nov. However, we do not know where these spots were positioned. However, Carré did remark that his nectophores measured up to 7 mm in length and 6 mm in width, which was considerably larger than the 3 mm in length and 3.2 mm in breadth that presently was found for the nectophores of C. ordinatum . The nectophores of C. isocarrei sp. nov. were somewhat larger, that is 5.5 and 3.9 mm respectively, but still notably less than Carré's measurements, for which there is no obvious explanation.
Fortunately, there are several excellent photographs of the siphosomal stem and the various zooids attached to it, and representative ones have been selected here. Figure 50 View FIGURE 50 shows the same arrangement of the cormidia, as was found for Cordagalma ordinatum , with, anterior to each gastrozooid, a palpon, some male gonophores and then some female gonophores. All the bracts had been detached. However, it is clear that the palpons were not attached basally and, although difficult to see in this photograph, the proboscis, at least the one to the lower left, has a basolateral attachment.
It can be seen that, in life, the gastrozooid was a long narrow tube with the proboscis region having a very narrow canal running through it. The basigaster was separated from the stomach by a constriction and was relatively long, particularly in comparison with the living ones of Cordagalma ordinatum (see Figure 10 View FIGURE 10 D), and the preserved ones of C. isocarrei sp. nov. ( Figure 44 View FIGURE 44 ). The gastrozooid appeared to be attached directly to the stem. The arrangement of the bracts on the siphosome is shown in Figure 51 View FIGURE 51 , and there appeared to be six per cormidium, although it was not clear if they were actually associated with any of the zooids.
The bracts shown in Carré's (1968, Plate II, fig. 2) look very similar to the ones in Figures 51 View FIGURE 51 & 52 View FIGURE 52 and so probably belong to Carré's other species (= Cordagalma isocarrei sp. nov.). They differed from the ones shown in Plate II, figure 1 and so one might presume that those belonged to C. ordinatum . This would also be true for the ones shown in Plate II, figure 4, as a small palpon attached basally by a short pedicle was shown. The only individual photograph of a bract in Carré's work (his Plate III, fig. 1) showed an extremely young bract. From Figure 52 View FIGURE 52 (left), it can be seen that the mature bracts of Carré's variant species have a very distinctive shape that has all the characters described for the mature bracts of C. isocarrei sp. nov., particularly with regard to the truncated region on the upper surface, the swelling at the distal end of the bracteal canal, and the presence of nematocysts on the lower part of the median ridge.
The very young bracts that Carré (1968) illustrated ( Figure 52 View FIGURE 52 , bottom right) were very distinctive, and other photographs showed that they belong to Carré's variant species, i.e. Cordagalma isocarrei sp. nov. The smallest bracts that have been found with the type specimen of C. isocarrei sp. nov. ( Figure 53 View FIGURE 53 ) only superficially resembled them, although the smallest, which was about the same size as Carré's ones, did show some resemblance, especially in the thickening of the bracteal canal and its distal swelling. The nematocysts that can clearly be seen on the median ridges were identified by Carré (1968) as stenoteles, with a capsule size of 8.2 x 7.5 µm. On the same bracts, and so presumably belonging to C. isocarrei sp. nov. he described the presence of atrichous isorhizas on all the other edges, but these are not evident in the photographs, and no sizes were given. It is probable that Carré's bracts were photographed before preservation and that the clear-cut arrangement of the ridges changed after preservation.
With regard to the tentilla, Carré (1968, Plate III, fig. 7) illustrated a tentillum without a "rostre". A similar photograph is shown in Figure 54 View FIGURE 54 . In comparison with that of Cordagalma ordinatum (see above) the difference is very striking, and the similarities with that of C. isocarrei sp. nov. (cf. Figure 46) are very striking. It is somewhat surprising that neither Claude Carré (1968), nor Danielle Carré (1973) who studied the development of Cordagalma ordinatum , made any reference to the developing tentilla ( Figure 55 View FIGURE 55 ), although Claude Carré took several photographs of them. Large numbers of these can be present on mature tentacles and often they were larger than the mature ones. Basically, they were more elongate than the mature ones with the stenotele nematocysts isolated at the proximal end, while the cnidocils at the distal end were only beginning to develop.
As noted above the palpons of Cordagalma isocarrei sp. nov. were mostly attached baso-laterally, with the proboscis region projecting out at an angle of c. 135°. Although Carré (1968) noted that amongst his variable characters there was the fact that some palpons were attached laterally rather than basally, he also noted that the intermediate, baso-lateral, attachment position could be found. One, if not both, of the palpons shown in Figure 50 View FIGURE 50 are certainly attached baso-laterally and of those shown in Figure 56 View FIGURE 56 most appear to be attached baso-laterally and, interestingly, a small oil droplet often can be seen on the side opposite to the proboscis. The one on the extreme right also showed how the proboscis region can be deformed, as was noted for several of the C. isocarrei sp. nov. palpons. The one next to it is the only one that appears to be attached laterally. Thus there is a great similarity between the palpons of Carré's other species and those of C. isocarrei sp. nov. that is further evidence that these are one and the same species.
Finally, several of Carré's photographs show parts of the siphosomal stem with moderately well-developed gonophores ( Figure 57 View FIGURE 57 ). It can be seen that the larger male gonophores, with their distinctive spadix, were borne on long narrow pedicles, while the female gonophores had shorter ones. The presence of anastomosing radial canals on the female gonophores was somewhat similar that that drawn by Haeckel (1888b, Plate XV, fig. 15), but it probably is not a specific character.
All in all, there seems little to doubt that the species, Cordagalma isocarrei sp. nov., described above is the same as the one that Carré (1968) inadvertently included in his 1968 redescription of C. ordinatum . The general morphology of the bracts is very similar and the more often than not baso-lateral attachment of the palpons of Carré's second species supports this conclusion.
Distribution. The six known specimens of Cordagalma isocarrei sp. nov. come from a very limited area in Monterey Bay, California, mostly concentrate around the so-called "Midwater site". It is probable that other observations have been made of this species, but the transparency and fragility of the specimens make in situ specific identifications almost impossible. Three of the specimens came from a depth of c. 200 m, while one was collected much shallower, at 115 m. However, the other two were much deeper, with the deepest at 421 m. The depth range, therefore, overlaps that of Cordagalma bimaculatum sp. nov.
Carré's (1968) other species appears to have come from a net sample taken at an unknown depth, in the vicinity of Villefranche-sur-Mer, France, in the northwestern Mediterranean Sea.
Etymology. The specific name, isocarrei , refers to the fact that the species found in Monterey Bay so closely resembles the second one that Carré mistakenly described as Cordagalma ordinatum that there can be little doubt that they are one and the same.













No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Physonectae |
|
Family |
|
|
Genus |