
Eleutherodactylus juanariveroi, Rios-López, Neftalí & Thomas, Richard, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.177263 |
|
DOI |
https://doi.org/10.5281/zenodo.5678561 |
|
persistent identifier |
https://treatment.plazi.org/id/03BD2622-9050-FF8A-FF71-14CAEB81F872 |
|
treatment provided by |
Plazi |
|
scientific name |
Eleutherodactylus juanariveroi |
| status |
sp. nov. |
Eleutherodactylus juanariveroi View in CoL , n. sp.
( Fig. 2 View FIGURE 2 ; Table 1)
Holotype: KU 306997, an adult female collected at Sabana Seca, Toa Baja Municipality ( Fig. 2 View FIGURE 2 ), in a seasonally flooded herbaceous wetland in the vicinity of the US Naval Security Group Activity Sabana Seca (USNS- GASS) and the Caribbean Primate Research Center, Puerto Rico (18°26.127’N, 66°12.092’W), 10–20 m elevation, by N. Rios and R. Thomas on 2 August 2005.
Paratypes: 45 (26 females, 18 males, 1 juvenile). Females (KU 306998–99, USNMS 563626-28, UPRRP 6340–2, UPRRP 6343, UPRRP 6348–9, UPRRP 6351–4, UPRRP 6356–7, UPRRP 6359, UPRRP 6360–2, UPRRP 6365, UPRRP 6367–9, and RT 14535 [Richard Thomas, private collection]), males (KU 307000–02, USNM 563623–25, UPRRP 6344–7, UPRRP 6350, UPRRP 6355, UPRRP 6358, UPRRP 6363–4, UPRRP 6366, UPRRP 6370, and RT 14501), juvenile ( UPRRP 6371), all from the vicinity of the USNSGASS, the Caribbean Primate Research Center, and the public lands in the Toa Baja Municipality, 18°26.049’N, 66°12.209’W, 18°26.127’N, 66°12.092’W, N. Rios and R. Thomas, 30 July 2005, 2 August 2005, 3 August 2005, and 23 August 2005.
Diagnosis. A member of the West Indian subgenus Eleutherodactylus , auriculatus section, martinicensis series (sensu Hedges, 1989) having extensive dorsal skin glandularity (Fig. 3), minute vomerine teeth, a distinctive carpal element (see description), a unique high-pitched call, and a palustrine habitat. Males have an external single subgular vocal sac, absent in females; nuptial pads absent. Pupil horizontally elliptical with two thin, sharp vertical slits, black colored below and above the mid portion of pupil, resulting in a ‘+’, sometimes only the inferior slit is clearly visible resulting in a ‘T’ ( Fig. 2 View FIGURE 2 ). The digital disks are small and spatulate as opposed to widening abruptly from the base of the pad. The terminal phalanges are nearly T-shaped, clearly visible in finger III and Toe IV, but only with minute lateral projections of the terminal phalanges; rest of phalanges knobbed clearly visible in finger I of holotype (KU 306997), although terminal transverse groove across the tip of the digital pad visible in external view. Large carpal element (fused carpals 2+3) with pronounced ventral spine and emargination (the equivalent structure in E. gryllus being more solid and rounded with a minute spine). Eyelid tubercles absent; dorso-lateral folds absent. Ulnar tubercles absent; thenar tubercle elevated, elliptical, about the same size as pad in finger I; subarticular tubercles rounded; few minute supernumerary tubercles on proximal segments of fingers; a few minute centrally grouped supernumerary palmar tubercles; fingers lacking lateral fringes; relative length of fingers 1=2<4<3; disk of fingers small, spatulate not widening abruptly ( Fig. 4 View FIGURE 4 A). Heel tubercles few, small, rounded, elevated; tarsal fold and lateral fringes of the toes absent; metatarsal tubercle rounded; subarticular tubercles rounded and slightly larger than palmar; supernumerary plantar tubercles present on proximal segments of toes; sole with a few minute irregularly scattered plantar tubercles; relative lengths of toes 1<2<3=5<4 ( Fig. 4 View FIGURE 4 B). Finger webs and toe webs absent.
The closest relative of E. juanariveroi is E. gryllus with which it agrees in size and proportions (Fig. 3, Tables 1 and 2 View TABLE 2 ), skin glandularity and fundamental call note. However, E. juanariveroi is smaller than E. gryllus (SVL in mm: males, 14.7 females 15.8 vs. E. gryllus males 15.7 and females 16.4) ( Table 1 and 2 View TABLE 2 ). The nares are prominent, and a ridge connects them behind the snout tip, giving it a somewhat squared-off appearance (Fig. 3A). On the other hand, the nares are less prominent in E. gryllus ; the loreal surface is steeper on the average (65–79°) in E. juanariveroi compared with E. gryllus (51–67°); the loreal surface has a midindentation, which is not so pronounced in E. gryllus . The vomerine teeth are minute (2–3 teeth in UPRRP 6358) or absent (UPRRP 6341); the vomerine teeth of E. gryllus are small, but always visible (2–3 teeth in UPRRP 6373 and UPRRP 6374). The large carpal element (fused 2+3) is sculptured and emarginated ventrally, much more so than in E. gryllus . Glands in E. juanariveroi are clearly discernible on both external and internal examination (KU 306998) while glands in E. gryllus can only be verified by examining the inner surface of the skin (UPRRP 6372); glands are heavier and more extensive over the posterior angle of the head (supra-axillary gland, parotoid glands sensu Lynch [1971]), body (flanks-lumbar-inguinal) and hind leg (dorsal side of thighs, tibiae, and tarsi) in E. juanariveroi than in E. gryllus (Fig. 3B); gular glands prominent and more extensive in females of E. juanariveroi than in E. gryllus ( Fig. 5 View FIGURE 5 A); the hyoid plate is narrower in E. juanariveroi than in E. gryllus ( Fig. 5 View FIGURE 5 b); left lobe of liver short and rounded, significantly smaller than right lobe compared with E. gryllus ( Fig. 5 View FIGURE 5 C); the ventral skin is unpigmented or with a very light stippling of melanophores and is smooth to weakly areolate in E. juanariveroi (pigmented, slightly glandular, and with more prominent areolae in E. gryllus ). The relationship between tympanum width and inter-narial distance and between tibio-fibula length and tarsal length are significantly smaller than in E. gryllus (see beyond, Multivariate comparison of morphological ratios, for a detailed explanation on how we identified diagnostic morphological ratios and assessed the statistical significance).
Eleutherodactylus juanariveroi Eleutherodactylus gryllus
Males (N = 18) Females (N = 27) Males (N = 11) Females (N = 10)
SVL 12.9–16.0 (14.66 ± 0.74) 12.1–17.3 (15.76 ± 1.21) 14.0–17.0 (15.65 ± 0.89) 14.1–17.9 (16.41 ± 1.18) HW 5.1–6.1 (5.61 ± 0.23) 4.7–6.5 (5.98 ± 0.39) 5.4–6.5 (6.07 ± 0.33) 5.7–7.1 (6.39 ± 0.44) HL 5.6–6.4 (5.95 ± 0.22) 5.4–7.8 (6.33 ± 0.48) 5.8–9.3 (6.83 ± 0.88) 6.3–7.4 (7.03 ± 0.41) UEW 0.9–1.4 (1.21 ± 0.11) 1.0–1.7 (1.27 ± 0.14) 1.1–1.7 (1.38 ± 0.17) 1.2–1.5 (1.38 ± 0.10) IOD 1.7–2.0 (1.82 ± 0.08) 1.4–2.3 (1.97 ± 0.18) 1.7–2.2 (2.00 ± 0.20) 1.8–2.4 (2.17 ± 0.21) TYW 0.6–1.0 (0.81 ± 0.09) 0.5–1.2 (0.95 ± 0.14) 0.8–1.1 (1.00 ± 0.09) 0.8–1.3 (1.09 ± 0.13) IN 1.3–1.5 (1.43 ± 0.08) 1.2–1.7 (1.51 ± 0.10) 1.3–1.7 (1.51 ± 0.12) 1.4–1.7 (1.60 ± 0.10) EL 2.1–2.6 (2.27 ± 0.14) 2.0–2.8 (2.36 ± 0.19) 2.3–3.0 (2.60 ± 0.25) 2.3–3.0 (2.73 ± 0.24) EN 1.3–1.7 (1.52 ± 0.09) 1.4–2.2 (1.70 ± 0.15) 1.6–2.1 (1.78 ± 0.15) 1.7–2.1 (1.93 ± 0.12) FD III 0.6–0.7 (0.67 ± 0.05) 0.5–0.9 (0.73 ± 0.08) 0.6–1.0 (0.76 ± 0.11) 0.6–0.9 (0.77 ± 0.08) FDB III 0.4–0.5 (0.44 ± 0.04) 0.4–0.6 (0.45 ± 0.04) 0.4–0.5 (0.44 ± 0.04) 0.4–0.5 (0.43 ± 0.03) TD IV 0.5–0.8 (0.67 ± 0.07) 0.5–0.9 (0.74 ± 0.09) 0.6–1.0 (0.78 ± 0.12) 0.6–0.9 (0.77 ± 0.10) FL 5.7–7.3 (6.59 ± 0.42) 5.4–7.5 (6.99 ± 0.46) 6.4–7.6 (7.00 ± 0.44) 6.5–8.9 (7.71 ± 0.63) TiL 5.9–7.2 (6.45 ± 0.33) 5.5–7.6 (6.99 ± 0.51) 6.6–8.7 (7.26 ± 0.60) 6.7–8.6 (7.70 ± 0.56) TaL 3.5–4.0 (3.84 ± 0.14) 3.3–4.6 (4.10 ± 0.28) 3.9–5.5 (4.48 ± 0.46) 3.9–5.1 (4.71 ± 0.35) Males Females The dorsal coloration of Eleutherodactylus juanariveroi in life is yellow to yellowish brown ( Fig. 2 View FIGURE 2 B) with a light, longitudinal, reversed comma mark on each side, lateral to the dorsal pigmented zone, with the head of the comma lying over the supra-axillary (parotoid) gland; the dorsal pigmented zone itself bifurcates, each branch being broad and fading, usually defining part of the cephalic triangle and not forming the sharp, well-defined “tines” of E. gryllus ; the pattern is generally non-contrasting, versus a strongly contrasting dorsal pattern in E. gryllus that may include reversed parenthesis marks and occasionally a bold yellow mid-dorsal stripe ( Fig. 2 View FIGURE 2 B; Joglar, 1998, Figs. 4.11–4.12; Rivero, 1998, Pl. XXV). Eleutherodactylus juanariveroi can be distinguished from other Puerto Rican species by an overall yellowish, non-contrasting coloration with a reversed comma pattern and a broadly bifurcated mid-dorsal zone ( Figs. 2 View FIGURE 2 B and 6).
The marks that we call inverted commas in E. juanariveroi may be homologues of the inverted parenthesis marks of E. gryllus , but they could also be a modification of the entire lateral zone ( Fig. 6 View FIGURE 6 ). However, these inverted commas may be a modification of the basic pattern elements of related species. Thus, in order to describe the color pattern of E. juanariveroi we relate it to the patterns of the other small Puerto Rican species ( E. cochranae , E. eneidae , E. gryllus , and E. locustus ) by identifying similar pattern elements ( Fig. 6 View FIGURE 6 ): (1) a dark mid-dorsal zone extending from roughly the sacral region to the head, usually narrowing gradually along the trunk and expanding again near the head, thus having curved indentations on either side, (2) a pair of light, narrow reversed parenthesis marks that closely follow the indentation in the mid-dorsal zone, (3) a bifurcation of the mid-dorsal zone near the head, producing two dark, diverging extensions that approach the posteromedial edge of each eyelid (in E. juanariveroi , these are pale and broad), (4) a median head triangle, an apex of which projects caudad, fitting between the diverging extensions of the divided mid-dorsal zone, with the other two angles at each eye and side extending between the eyes (another pattern element that occurs in some of these species is a light midline stripe, but it is an occasional variant and is not found in E. juanariveroi ). These pattern elements have been variously modified by adding or lessening of pigment in the different species (and are variable within species), but reversed parentheses are frequent. A further modification occurs when pigment bridges form between the lower parts of the “tines” of the fork to the caudad apex of the cephalic triangle, which results in a dark scapular ‘W’ ( Fig. 6 View FIGURE 6 ).
On some individuals of these related small species, particularly E. locustus , the scapular ‘W’ and the light reversed parentheses may be the only pattern readily seen. Some of the components occur in frog species we have not mentioned; for example, E. hedricki (Rivero) and E. eneidae may have the reversed parentheses. In E. juanariveroi the pattern is much reduced in contrast; the reversed parenthesis marks are very wide; and the median dark zone is relatively light and the anterior branches broad; a well-developed scapular ‘W’ is not found. The result is a pair of light, longitudinal marks that curve laterally onto each side of the occiput, terminating with the large posterior head glands ( Figs. 2 View FIGURE 2 B and 6). Interior to these marks on many specimens is another pair of diverging, light longitudinal marks (the altered median dorsal zone and branches). Some individuals show only the light heads of the commas, the rest of the pattern being obscure; this results in a pattern of two ocelli over the area of the scapula on the back of the head ( Fig. 6 View FIGURE 6 ), representing the most divergent pattern among this group of small frogs.
A light-brown inter-orbital band that extends to the tip of the snout is present in 16.3% of the E. juanariveroi (clearly defined in KU 306999) and absent or rare in E. gryllus (some specimens are so faded that it is difficult to discern the pattern). The legs of E. juanariveroi do not have any cross bands; those of E. gryllus have 1–2 hazy bands on the thigh and two on the shank. Although its call is related to that of E. gryllus , it is distinctive in its patterning and higher pitch (see Advertisement call for a detailed explanation on call parameters).
A Puerto Rican Eleutherodactylus that may has morphological affinities with E. juanariveroi and E. gryllus is E. jasperi . Eleutherodactylus jasperi also has two pairs of lateral sub-dermal glands well developed on flanks and posterior surfaces of thighs ( Drewry and Jones, 1976). However, although E. jasperi has a yellow to yellow-brown body coloration in life, it differs dramatically from E. juanariveroi and E. gryllus in reproduction ( E. jasperi is ovoviviparous, the other species are oviparous), body size (19–20 mm in E. jasperi , 14– 15 mm in E. juanariveroi , 15–16 mm in E. gryllus ), acoustics (frequency of calls: 5 kHz in E. jasperi , ~7 kHz in E. gryllus , ~ 8 in E. juanariveroi ), habits ( E. jasperi is an obligate bromeliad-dwelling in somewhat xeric conditions in highlands, E. juanariveroi is an obligate herbaceous-dwelling in palustrine lowland wetland [N. Rios, in prep.], E. gryllus is arboreal in highland wetland conditions), and geographic distribution. Additionally, E. jasperi lacks prevomerine teeth while E. juanariveroi and E. gryllus have them.
Measurements (in mm) and description of the holotype. Adult female; SVL 16.7; head width 6.1; head length 6.7; upper eye lid width 1.1; inter-ocular distance 2.1; tympanum width 1.0; inter-narial distance 1.5; eye length 2.3; eye-tympanum distance 0.3; eye-naris distance 1.7; fingertip (III) width 0.8; toe tip (IV) width 0.9; femur length 7.5; tibia-fibula length 7.2; tarsal length 4.2. Inter-orbital band absent; gular glands prominent; supra-axillary gland (parotoid), body glands (flanks-lumbar-inguinal), and hind limb glands (dorsal side of thighs, tibiae, and tarsi) clearly visible; well-expressed pale reversed comma heads and a pale dorsal zone bifurcation occur in dorsum. Mature eggs undetected externally. Dorsal coloration in life yellow to light brown; light brown in preservative.
Variation. SVL of adult males 12.9–16.0 mm (mean = 14.66, n = 18), of adult females 12.1–17.3 mm (mean = 15.76, n = 27), and juvenile (SVL 5.82 mm) of moderate habitus, typical of scansorial species of Eleutherodactylus ( Table 1); head around 40% of SVL; snout tip pointed in dorsal view, sides forming an angle of about 72°; nares distinctly protuberant, canthus rostralis not distinct, lores sloping, slightly indented; eyes prominent, EL into HW around 2.7; tympanic membrane present and tympanic annulus distinct but with roughly the dorso-posterior third of the rim obscured by supra-tympanic fold, fitting tightly between the ventro-posterior edge of the orbit and the angulus oris, TYW into HW around 6; 2–3 minute vomerine teeth barely visible on weakly defined patches postero-medial to choanae (of two cleared and stained specimens UPRRP 6358, a male, with 3 tiny vomerine teeth on one patch, 2 or 3 in the other patch, teeth absent in UPRRP 6341, a female); tongue rounded posteriorly with a weak notch; there are well-developed glands on the following regions (Fig. 3): (1) supra-axillary (parotoid), extending ventrally to glenoid region, (2) flanks to sacral region, (3) posterior surfaces of thighs, shank, tarsi and (4) postero-lateral parts of the gula (sexually dimorphic: present in 96.3% in females, which also have larger glands, compared with 23.5% in males). Digital disks moderate, fitting within the tympanum; hyoid plate narrow ( Fig. 5 View FIGURE 5 B); medial element of anterior process of the hyale long and straight; hyale without free flanges; antero-lateral process angled anteriorly; postero-lateral process long and thin; thyrohyal process narrow. Venter nearly smooth with weak, rounded areolations and with a sparse stippling of melanophores; testes pigmented. Measurements (range, mean, and standard deviation) of the type series are given in Table 1 along with measurements of E. gryllus for comparative purposes.
The limbs are irregularly pigmented with varied intensities of pigment but without a distinct patterning; the post-femoral surfaces of some specimens are darker; the digit tips of manus and pes are darkly pigmented. In some individuals of E. juanariveroi , a horizontally elliptical pupil, and a thin, dark vertical slit below the mid portion of pupil result in a ‘T’ ( Fig. 2 View FIGURE 2 A) similar to the condition in E. gryllus , E. eneidae , and E. locustus . In other individuals of E. juanariveroi the pupil resembles that in E. cochranae , which has thin, vertical slits below and above pupil resulting in a ‘+’. Pupil shape in E. juanariveroi , however, differs from E. brittoni , which lacks vertical slits.
Multivariate comparison of morphological ratios. Non-metric Multidimensional Scaling (NMDS) of morphological ratios of adult specimens extracted two axes that explained 95.1% of the variance in ordination space (final stress = 11.3 and instability = 0.052 after 500 iterations, orthogonality = 94.8%) ( Fig. 7 View FIGURE 7 A). A Multi-Response Permutation Procedure (MRPP, a non-parametric analysis used to test the hypothesis of six species in the data matrix) revealed six groups that had similar within-group homogeneity (A = 0.27, p <0.0001), which corroborates the hypothesis of six nomenclaturally and taxonomically stable species in the data matrix. However, the overlap between E. juanariveroi and E. gryllus in ordination space suggests a strong affinity in morphological ratios between species. An Indicator Species Analysis (ISA) ( Legendre & Legendre, 1998) that could be helpful in distinguishing E. juanariveroi from other species resulted in four potential diagnostic morphological ratios (morphological ratio and Indicator Value from Monte Carlo tests with 10,000 permutations): SVL:HW, 17.1 (p = 0.067); TYD: IND, 18.9 (p = 0.004); TYD:EN, 18.2 (p = 0.028); TiL:TaL, 17.1 (p = 0.054). In ordination space, only TYD:IN and TiL:TaL differed significantly between males of E. juanariveroi and the most similar E. gryllus ( Table 2 View TABLE 2 ), which justified the use of these two morphological ratios to distinguish E. juanariveroi from E. gryllus ( Figs. 7 View FIGURE 7 B and 7C).
Distribution and natural history. Eleutherodactylus juanariveroi is known only from the type locality ( Fig. 1 View FIGURE 1 ), which lies within the subtropical moist forest life zone ( Ewel & Whitmore, 1973). This locality consists of a palustrine herbaceous wetland at 17 m elevation. The wetland is seasonally flooded with fresh water; the soil consist of swamp and marsh organic deposits from Pleistocene and/or recent origin ( Briggs & Akers, 1965). Herbaceous vegetation in this habitat consists of the toothed midsorus fern ( Blechnum serrulatum ), willdenow’s maiden fern ( Thelypteris interrupta ), bulltongue arrowhead ( Sagittaria lancifolia ), flatsedges ( Cyperus sp.), spike rushes ( Eleocharis sp.), and vines and grasses. The habitat at the type locality occupies approximately180 ha; we were unable to find the frog at other wetland sites in the northern coastal plain. All specimens were collected between 1900–2200 h while perching, sitting, or calling on herbaceous vegetation, mainly on ferns, between 0.4 m and 1.2 m above water level. Calling sites for males were 0.6 to 1.2 m above the ground. Egg clutches comprised 1– 5 eggs and were found on leaf axils (21 egg clutches) or leaf surfaces (3 egg clutches) of only S. lancifolia (N. Rios, unpubl. data). Like E. gryllus , the calling activity of E. juanariveroi started at approximately 1630 h and decreased significantly before midnight. The single locality from which E. juanariveroi is known is adjacent to the coast at 17 m elevation, while E. gryllus is only known from between 305 m to 1189 m elevation ( Joglar, 1998), although we have not recently found E. gryllus below 600 m elevation. This species is mostly arboreal, calling from branches and bromeliads, which contrasts with E. juanariveroi .
Advertisement call. The call consists of a series of short high-pitched notes ( Fig. 8 View FIGURE 8 A). At 26 C, call duration varies from 4–21 sec (10.9 ± 6.2 sec, n = 6). Call structure consists of one to three one-note series, followed by one or more series of two notes each, one (or more) series of three-four notes each, and a series that varies from 5–30 notes at the end of the call. Notes per call was 8–45 (23.0 ± 13.4 notes, n = 6) ( Fig. 8 View FIGURE 8 A). Frequency was 6.03–9.02 kHz; the dominant frequency was 7.38–8.28 kHz (7.69 ± 0.18 kHz, n = 92) ( Fig. 8 View FIGURE 8 A). Note duration was 15–31 ms (24.1 ± 3.1 ms, n = 92). Note repetition rate varies between consecutive notes within a series of two or more notes (341 ± 22 ms, n = 58) and between the final note of a series and the note in the following series (819 ± 99 ms, n = 30) ( Fig. 8 View FIGURE 8 A). The advertisement call of E. juanariveroi differs from that of E. gryllus in notes per call (range 5–29, 15.3 ± 10.0 notes, n = 4) ( Fig. 8 View FIGURE 8 B), dominant frequency (range 6.31–7.79 kHz, 7.44 ± 0.28 kHz, n = 61) ( Fig. 8 View FIGURE 8 B), note duration (range 21–32, 26.8 ± 2.4 ms, n = 61), and note repetition rate (consecutive notes within a series 1.23 ± 0.13 sec, n = 53; between the first note of the call and the note in the following series in the call 1.97 ± 0.67 sec, n = 4) ( Fig. 8 View FIGURE 8 B). Among sympatric Eleutherodactylus , the call of E. juanariveroi has the highest frequency and the greatest structural complexity compared with E. brittoni , E. cochranae , and E. coqui (Thomas) , but probably among all Puerto Rican Eleutherodactylus as well (for comparison see Drewry & Rand, 1983 and Narins, 1995).
Etymology. The specific name honors Juan A. Rivero, distinguished Puerto Rican herpetologist and naturalist.
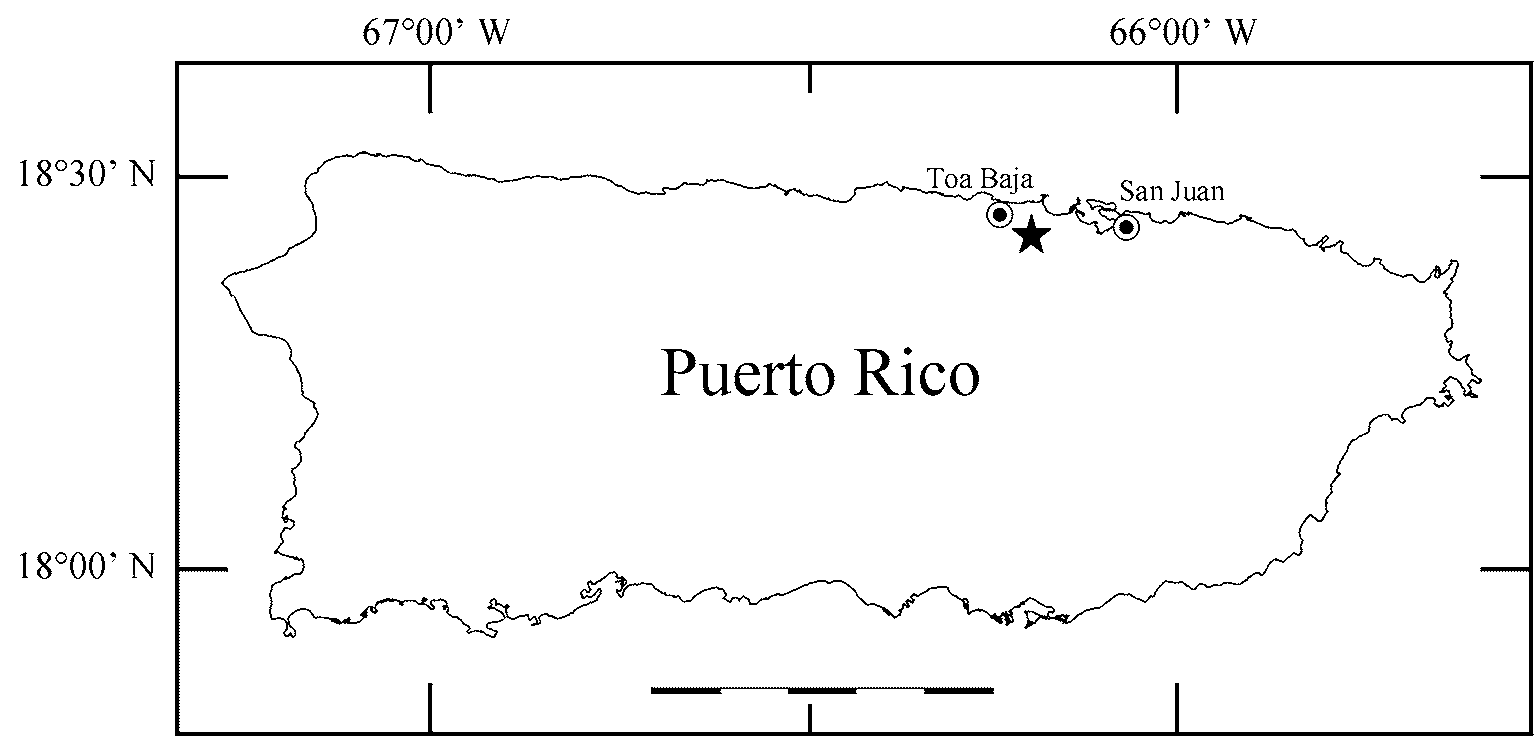

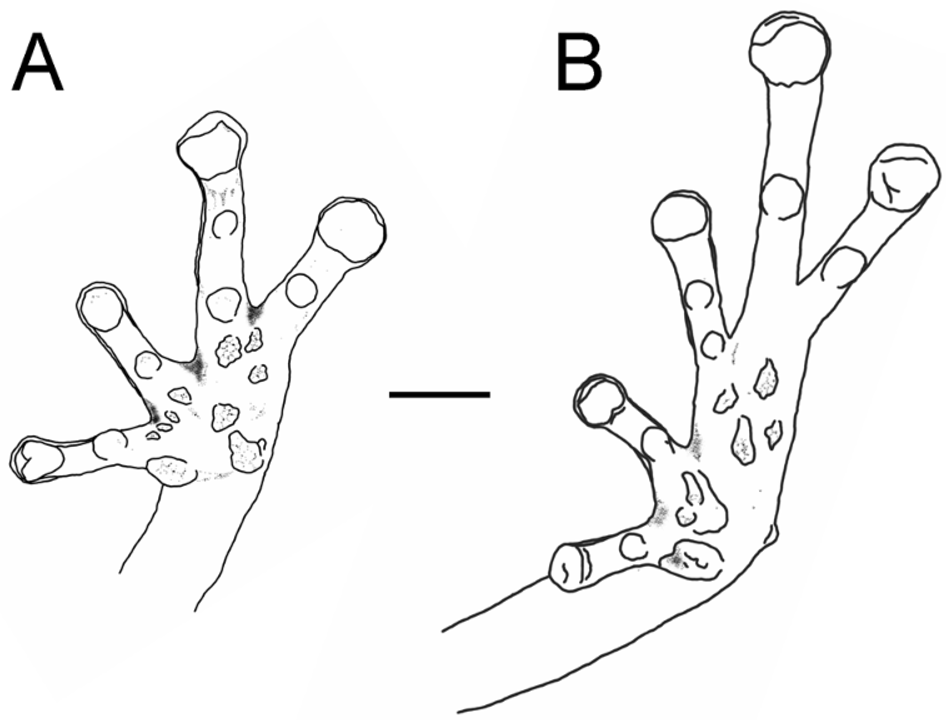
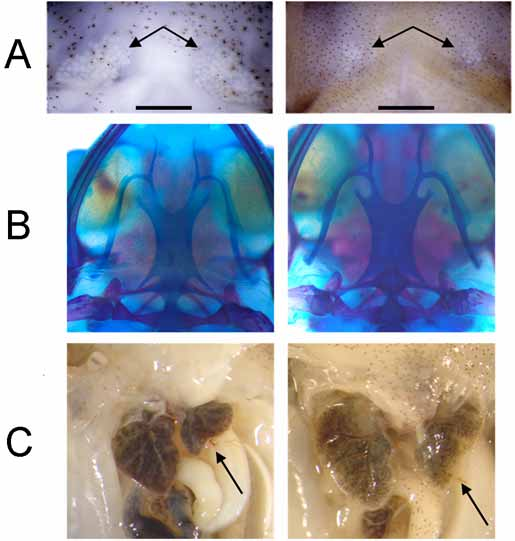
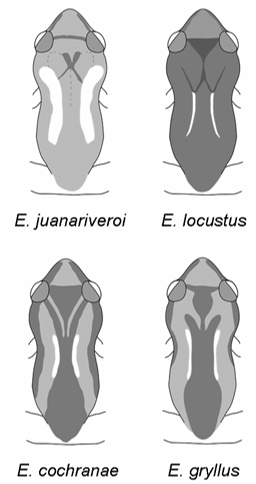
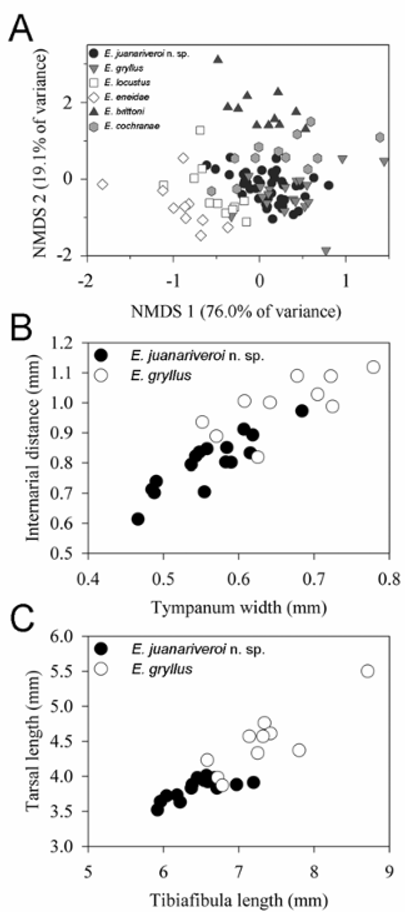
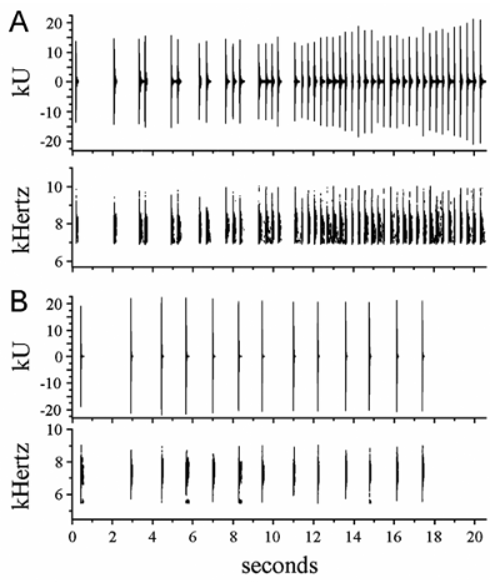
TABLE 2. Body proportions (mean ± SD) of Eleutherodactylus juanariveroi and Eleutherodactylus gryllus that showed significant differences at α = 0.05 and α = 0.10. Significant variables (TYW / IN and TiL / TaL) were extracted from NMDS multivariate analysis. Data for males and females were analyzed separately; Mann-Whitney U-Test.
| E. juanariveroi (N = 18) | E. gryllus (N = 10) | Z | P | E. juanariveroi E. gryllus (N = 27) (N = 9) | Z | P | |
|---|---|---|---|---|---|---|---|
| TYW/IN | 0.56 ± 0.06 | 0.66 ± 0.07 | 3.03 | <0.01 | 0.63 ± 0.07 0.68 ± 0.06 | 1.60 | 0.11 |
| TiL/TaL | 1.68 ± 0.06 | 1.64 ± 0.09 | 1.66 | 0.09 | 1.70 ± 0.06 1.65 ± 0.07 | 1.34 | 0.18 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.