
Cardiodactylus enkraussi Otte, 2007
|
publication ID |
https://doi.org/ 10.5252/z2009n3a11 |
|
persistent identifier |
https://treatment.plazi.org/id/03BDFA31-9D02-FF94-3D87-FBCDFCF2B1BE |
|
treatment provided by |
Marcus |
|
scientific name |
Cardiodactylus enkraussi Otte, 2007 |
| status |
|
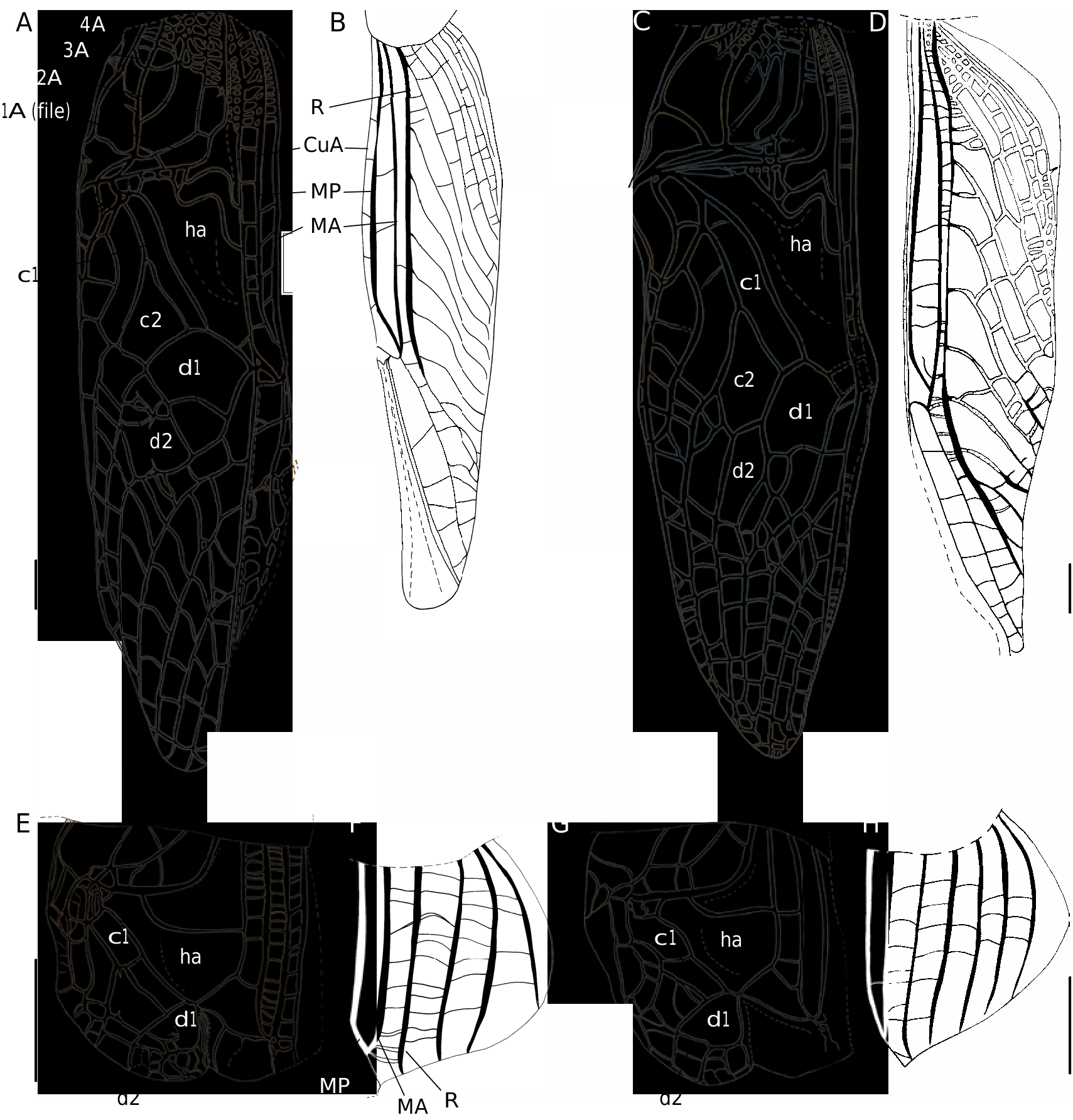
Cardiodactylus enkraussi Otte, 2007 View in CoL ( Figs 1A, B View FIG ; 2 View FIG ; 3C View FIG ; 4 View FIG A-C;
5A; 6A; 7A-C; 14; 17A-C)
Cardiodactylus enkraussi Otte, 2007a: 353 View in CoL ; 2007b: 32. — Robillard et al. 2007: 1254.
TYPE MATERIAL. — Vanuatu. [Sanma Province], [Espiritu] Santo [Is.], Segond Channel, VIII.1950, Krauss, ♂ holotype ( BPBM 16,789). — Paratype ♀: identified by Otte (2007a) without specification. Not examined.
TYPE LOCALITY. — Vanuatu, Espiritu Santo Is., Segond Channel.
MATERIAL EXAMINED. — Vanuatu (46 ♂♂, 38 ♀♀, 23 juv.). [Province Sanma], Espiritu Santo Is., Luganville, site du CTRAV, forêt côtière sur sable, 15°27’00’’S, 167°12’26’’E, 28.X.2006, nuit, 1 ♀ (TR-409), sur plante h = 1 m GoogleMaps ; 31.X.2006, nuit, 1 ♂ (TR-482), sur plante, photo S. Hugel, PIII: molec, T. Robillard (MNHN-EN- SIF2312, 2313). — Nattawa, forêt pâturée, 15°19’29’’S, 167°12’09’’E, 29.X.2006, jour, 2 ♀♀ (TR-428-429), sur plante; 1 ♂ (TR-431), sous feuille de plante de sous-bois, T. Robillard (MNHN-ENSIF2314-2316). — Peavot , rive N de la rivière principale, terrasse I, 14°59’37’’S, 166°47’04’’E, 38 m alt., forêt secondaire (ancien jardin), 20.X.2006, jour, 9AM-3PM, 1 ♀ (TR-247), litière et herbes basses, “repos” puis saute et vole si dérangée, 1 ♂ juv. (TR-238), 1 ♀ (TR-239), 1 ♀ juv., sur plante, 21.X.2006, nuit, 3 ♂♂ (TR-293-295), 1 ♀ (TR-289), sur plante, 1 ♂ (TR-331), sur plante de sous-bois h = 0,3 m, enregistrement en milieu naturel TR-take 331 GoogleMaps ; 25.X.2006, nuit, 1 ♂ (TR-331), sur plante de sous-bois h = 0,3 m, enregistrement en milieu naturel TR-take 331, T. Robillard (MNHN-ENSIF2317-2326); 20.X.2006, jour, 4 juv. (nº 46, 48, 71, 72), litière; 21.X.2006, nuit, 2 ♂♂ (nº 60, 61), 1 ♀ (nº 63), 1 juv. (nº 64), sur plante, 1 ♀ (nº 62), sur plante, PIII molec, 1 ♂ juv. (nº 79), sur plante de sous-bois h = 0,3 m, sur feuille mangeant une fleur, tube 5-2006, L. Desutter-Grandcolas ( MNHN- ENSIF2327-2336 ). — Peavot, Mont Nouresororo, 14°59’26’’S, 166°45’14’’E, 636 m alt., forêt de brume, 22.X.2006, nuit, 1 ♂ (TR-298), sur plante de sous-bois h = 0,5 m, en train de manger sur feuille, photo TR, 1 ♂ (TR-299), sur plante, enregistrement en semicaptivité (chant d’appel) TR-take 64-67, T. Robillard (MNHN-ENSIF2337, 2338) GoogleMaps ; 22.X.2006, nuit, 1 ♀ juv. (nº 6), sur plante de sous-bois, L. Desutter-Grandcolas (MNHN-ENSIF2339). — Peavot, rive S de la rivière principale, terrasse I, 14°59’37’’S, 166°47’04’’E, 38 m alt., forêt secondaire (ancien jardin), 20.X.2006, jour, 9AM-3PM, 2 juv. (TR-230, 231), litière, 1 juv. (TR- 228), sur plante, photo TR, 1 ♀ (TR-248), sur tronc, “repos” puis saute et vole quand dérangé; 20.X.2006, jour, 1 ♂ (TR-245), sur plante h = 0,4 m sous les feuilles, 1 ♂ (TR-246), litière, inactif; 21.X.2006, nuit, 2 juv. (TR-281, 282), sur plante; 24.X.2006, nuit, 1 ♀ (TR-310, alcool), PIII: molec, T. Robillard ( MNHN- ENSIF2340-2348 ) ; 21.X.2006, jour, 1 ♀ juv. (nº 9), sous écorce détachée, arbre à contrefort, h = 1,20 m, photo, L. Desutter-Grandcolas (MNHN-ENSIF2349). — Peavot, rive S de la rivière principale, terrasse II, 14°59’37’’S, 166°47’04’’E, 38 m alt., forêt secondaire (ancien jardin), 23.X.2006, nuit, 1 ♀ (nº 16), sur arbre, L. Desutter- Grandcolas (MNHN-ENSIF2350). — Butmas, forêt du plateau de Tankara, 15°21’56’’S, 166°59’E, 600 m alt., 16.X.2006, jour 10AM, espace dégagé près du camp, herbes hautes en bord de piste h = 1 m, 1 ♂ (TR-119), photo SH, 1 ♀ (TR-117), 1 ♀ (TR-118), photo SH; 16.X.2006, nuit 6-10PM, 1 ♀ (TR-133), sur plante de sous-bois h = 2 m, pied de falaise, photo site LDG, PIII molec, 1 ♂ (TR-126), sur plante; 17.X.2006, nuit, 1 ♂ (TR-148), sur plante de sous-bois h = 1 m, bord de chemin, 1 ♂ (TR-147), herbes hautes en bord de piste/lisière, sur branche fine dans la végétation, T. Robillard (MNHN-ENSIF2351-2357). — Grotte de Fapon /sentier, 15°19’51,6’’S, 166°57’53,6’’E, forêt dans la doline, 14.X.2006, nuit, 1 ♂ (TR-76), sur plante de sous-bois h = 1,5 m en bord de piste près du camp, sous une fougère, enregistrement en semi-captivité (camp) takes 23, 25 GoogleMaps ; 15.X.2006, jour, 1 ♂ (TR-80), adulte en élevage, pelouse/zone herbacée en bord de sentier, 1 ♀ (TR-84), sur feuille de plante de sous-bois, T. Robillard (MNHN-ENSIF2358-2360). — Matantas, Big Bay, Vatthé Conservation area , 15°20’S, 166°95’E, 25.X.2006, nuit, 2 ♀♀ (TR-328, 329), sur plante, 1 ♂ (TR-330), sur plante de sous-bois h = 0,6 m, sur feuille en arrière plan, enregistrement en milieu naturel TR-take 62 ; 27.X.2006, jour, 1 ♀ (TR-363), repos dans litière, photo TR-82-86, 1 ♀ juv. (TR-369), sur plante; X.2006, jour, 1 ♂ (TR-346, alccol), PIII: molec, T. Robillard (MNHN-ENSIF2361-2366). — Big Bay, Vatthé Conservation area , 15°20’S, 166°95’E, 26.X.2006, nuit, 3 ♂♂ (nº 46, 47, 48), 1 ♀ juv. (nº 49), sur plante de sous-bois ; 27.X.2006, nuit, 3 ♂♂ (nº 52-54), sur plante de sous-bois, L. Desutter-Grandcolas ( MNHN- ENSIF2367-2373 ). — Big Bay, Vatthé Conservation area , forêt secondaire près d’une grotte à chauve-souris: 26.X.2006, jour, 1 ♀ (TR-354), sur plante, photo habitat TR, 1 ♂ (TR-353), 1 juv. (TR-355), 2 ♀♀ (TR-348, 350), sur plante, 1 ♂ (TR-351), sur tige ; 27.X.2006, nuit, 1 ♂ (TR-384) photo SH – 2 autres individus sur la même feuille: échant. bot. 4, 2 ♀♀ (TR-386), 2 ♂♂ (TR-390, 391), 1 ♀ juv. (TR-392), sur plante, T. Robillard (MNHN-ENSIF2374-2384). — Nattawa , forêt pâturée, 15°19’29’’S, 167°12’09’’E, 29.X.2006, jour, 2 ♀♀ juv. (TR-422, 433), sur plante, T. Robillard (MNHN-ENSIF2385, 2386). — Funafus, 15°32’46,4’’S, 167°00’46’’E, 30.X.2006, jour, 1 juv. (TR-460), 1 ♀ juv. (TR-461), 1 ♀ (TR-463), sur plante, 2 ♀♀ (TR-454, 455), sur plante h = 0,4 m, comportement de fuite: saute et vole si dérangée, T. Robillard (MNHN-ENSIF2387- 2391). — No precision, X.2006, 4 ♀♀, 2 ♂♂, T. Robillard (MNHN-ENSIF2392-2397). — Penaoru, Malaise, sol, 900 m A., 6-18.XI.2006, MG9 A1, 1 ♀ (MNHN-ENSIF2398), Malaise, sol, 900 m C., 6-18. XI.2006, MG9 C1, 1 ♀ C. Villemant (MNHN-EN- SIF2399). — Penaoru, 600 m, XI.2006, 2 ♂♂ (ortho 14), Exped. SANTO 2006 (MNHN-ENSIF2400, 2401). — Reared in laboratory: F1, 2006-2007, 6 ♀♀, 9 ♂♂ (6 recorded, MNHN-ENSIF2402-2407), 9 juv. GoogleMaps ; F2, 2007-2008, 1 ♂ (recorded, MNHN-ENSIF2408), T. Robillard. — [Vanuatu] Nouvelles-Hébrides. [Province Malampa], Malekula Is., 1 ♀, identified C. canotus by T. Robillard (Robillard & Desutter-Grandcolas 2004a, 2006), E. Aubert de la Rüe (MNHN-ENSIF1178) .
DISTRIBUTION. — Vanuatu, Espiritu Santo Is. and Malekula Is. Otte (2007a) also mentions specimens from Eromango Is. but none could be examined in the present study.
DIAGNOSIS. — Size rather small, body slender, general colouration dark brown to grey brown, quite homogeneous with a characteristic two-part calling song with low intensity compared to that of C. novaeguineae and C. tankara n. sp. Species very similar to C. pentecotensis n. sp. (see below), but differing by male genitalia (endophallic sclerite).
REDESCRIPTION
In addition to the characters given in the original description: size small, body slender, general colouration dark brown to grey brown, quite homogeneous ( Figs 2 View FIG ; 17B, C View FIG ). Scapes and antennae generally homogeneously yellowish brown. Pronotum: dorsal disk trapezoidal, bisinuated posteriorly. Legs yellowish brown, femora more or less mottled with brown; tibiae more or less annulated. Fore tibiae bulbous at tympana level. Hind tibiae dorsal face with 7-11 inner (m = 9, n = 10) and 11-17 outer (m = 15, n = 10) spines above spurs, and 6-9 inner (m = 8, n = 10) and 8-11 outer (m = 9, n = 10) spines between spurs. Tarsomere III-1 with 3-7 spines on dorso-external edges (m = 4, n = 10). Hind wings longer than FWs, forming a dark brown tail four times as long as pronotum, increasing species gracile shape.
Male
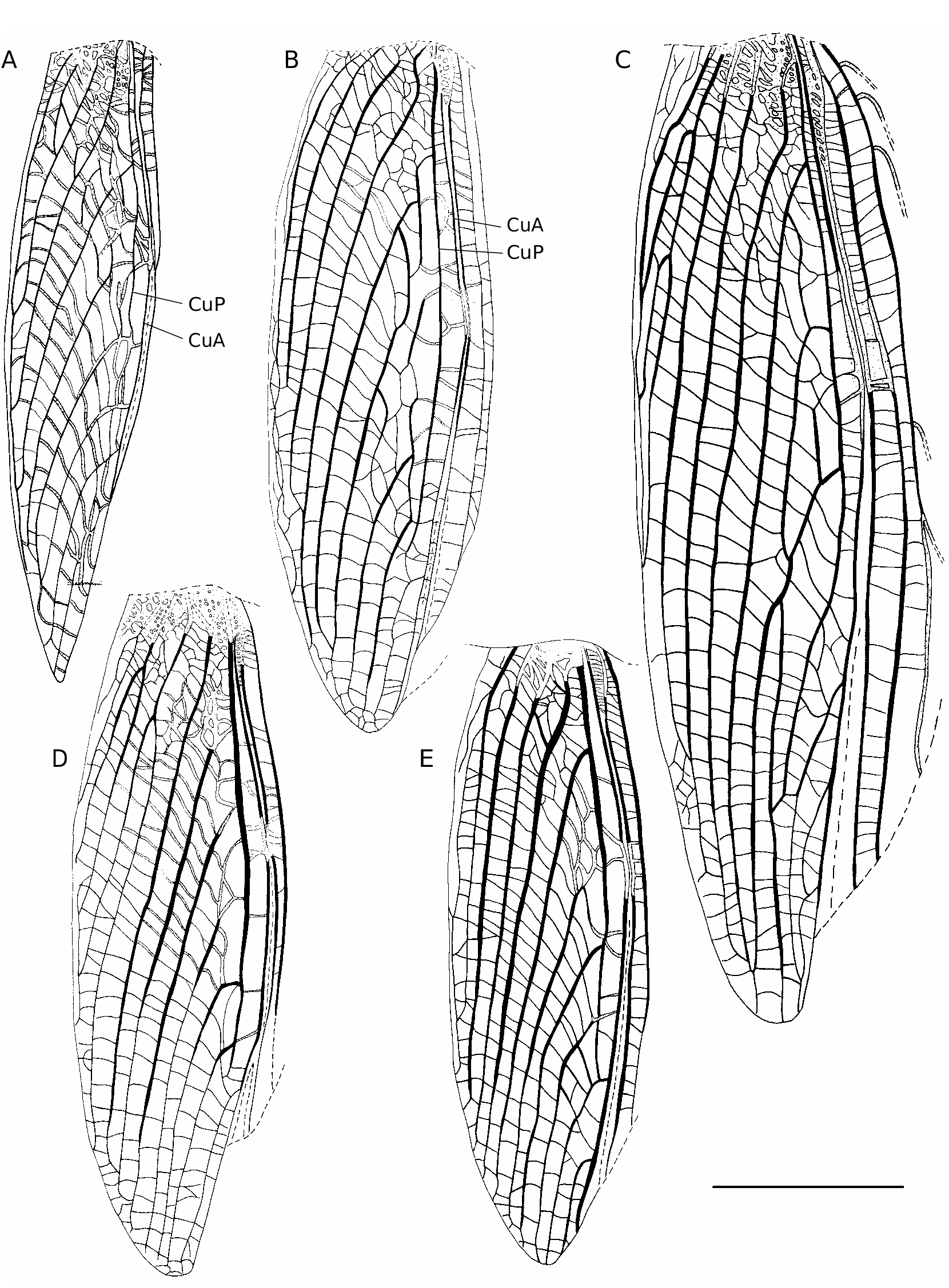
FW ( Fig. 1A, B View FIG ). Dorsal field grey brown, translucent, quite homogeneous; veins whitish to grey brown; with faint dark spots, sometimes absent: above and along 1A, on FW inner edge anterior to 1A, in cell e1, in middle of harp near harp veins. Lateral field with sometimes a dark band on the dorsal edge, including veins MA, MP and R; ventral edge lighter. FW venation close to that of C. novaeguineae . 1A bisinuated ( Figs 1A View FIG ; 3C View FIG ); stridulatory file with 166-179 teeth (m = 172, n = 5), located on both transverse (114-127 teeth, m = 119) and basal longitudinal (44-65 teeth, m = 52) parts of 1A ( Fig. 3C View FIG ). CuP missing. Harp narrow, 2 W-shaped harp veins. Mirror area variable, c1 long and narrow, c2 quite large; mirror (d1) prolonged by d2, forming together an ellipsoid surface, variably crossed by one or several accessory veins. Apical field lanceolate, with 4-5 longitudinal veins. Lateral field: latero-dorsal angle made by MP; R with 6-9 bifurcating veins.
Male genitalia ( Fig. 4 View FIG A-C): pseudepiphallus narrowed at midlength, with a large dorsal concavity and a narrow median process, slightly setose basally, concave dorsally, its apex spoon-like and more or less rounded. Pseudepiphallic parameres trilobate, a narrow divergent lobe fused with lateral membrane and two inner ventral lobes of variable shape. Membrane between base of parameres and ectophallic arc with a sclerite oriented posteriorly, forming wide membranous structures. Ectophallic arc complete and straight, anterior to parameres, with a median posterior expansion. Ectophallic apodemes variable in shape, not reaching rami apex. Apex of ectophallic fold between pseudepiphallic parameres, trilobate, the median lobe sponge-like, possibly glandular. Endophallic sclerite small, V-shaped, with a medio-posterior expansion. Endophallic apodeme made of a large medio-dorsal crest extended anteriorly. Membrane of endophallic cavity finely and transversally plicate.
Female
FWs grey brown to dark brown ( Fig. 17C View FIG ), veins slightly lighter than cells or of same colour, with lighter spots along CuA. FW venation ( Fig. 5A View FIG ): 9 or 10 strong longitudinal veins on dorsal field (3 anal veins). Ovipositor: shorter than hind femora; dorsal edge of apex slightly denticulate ( Fig. 6A View FIG ). Female genitalia ( Fig. 7 View FIG A-C): copulatory papilla rounded with a basal sclerotized ring; apex narrowed and folded ventrally, against a rounded ventral bump.
Juvenile
Colouration variable from yellowish brown to dark brown, dorsum and femora more or less mottled with dark brown, tibiae with dark bands; lateral sides and sternites black, except a median yellow stripe. Head dorsum with same colouration as in adults; face dark brown to black with yellow patterns.
Measurements
See Table 3.
HABITAT AND LIFE HISTORY TRAITS
In Espiritu Santo, C. enkraussi lives in forested areas ( Fig. 17A View FIG ) together with C. tankara n. sp. (Butmas area) or C. novaeguineae (coastal areas), although in less conspicuous places.
Individuals are found deep in vegetation, on between large leaves, on stems, bushes or low trees, between 30 cm and 3 m in height. Th e species is active at night only, but some specimens are found in leaf litter by day, from where they escape by jumping and flying. At night, groups of specimens gather on plants in relatively dense populations ( Tables 1; 2).
BEHAVIOUR
Calling song ( Fig. 14 View FIG ; Table 4)
Males call from dusk to late at night, staying deep within vegetation, on leaves or branches. The calling song of C. enkraussi is less conspicuous than that of C. novaeguineae and C. tankara n. sp. The songs of five males recorded in the field (TR-330, TR-331, MNHN-ENSIF2363, 2325) and in the laboratory (spec-El-3 [MNHN-ENSIF2404], El-4 [2405], El-7) at 22-26.5°C are analysed here. At 25°C, the call consists of irregular song phrases (mean phrase duration ± SD = 28.9 ± 2.6 s). Each phrase generally begins with a series of 28 ± 10, low amplitude, single echemes, followed by a series of 22 ± 2 more intense double echemes subdivided into two sub-echemes by an interval of 174 ± 23 ms, and/or by 1 to 3 intermediate syllables (m = 2.13 ± 0.45) sometimes joining the two sub-echemes.
The initial single echemes (SE) have the following characteristics: SE rate = 103.2/min; SE duration = 169 ± 27 ms; SE period = 581 ± 232 ms; SE duty cycle = 29%. Each SE includes 8.9 ± 1.3 syllables with the following characteristics: SE syllable rate = 56.5 ± 3.2/s; SE syllable duration = 6.6 ± 1.4 ms; SE syllable period = 20.5 ± 9 ms; SE syllable duty cycle = 32%.
The double echemes (DE) have the following characteristics: DE rate = 66/min; DE duration = 429 ± 27 ms; DE period = 909 ± 120 ms; DE duty cycle = 47%; DE total number of syllables = 21.7 ± 1.7. Th e first sub-echeme (DE1) of the double echeme is a low amplitude echeme equivalent to the initial single echemes. Each first sub-echeme includes 7.9 ± 1.1 syllables with the following characteristics: DE1 syllable rate = 60.5 ± 3.5/s; DE1 syllable duration = 5.9 ± 1.2 ms; DE1 syllable period = 18.2 ± 2.6 ms; DE1 syllable duty cycle = 32%. The second sub-echeme (DE2) of the double echeme has much higher amplitude and includes 11.9 ± 1.3 syllables with the following characteristics: DE2 syllable rate = 90.2 ± 5.0/s; DE2 syllable duration = 7.2 ± 2.4 ms; DE2 syllable period = 13.1 ± 1.2 ms; DE2 syllable duty cycle = 55%.
The dominant frequency of C. enkraussi is the third harmonic of the song (single echeme: 19.5 ± 0.6 kHz) (Robillard et al. 2007). Within the double echemes, the dominant frequency is slightly higher in syllables of DE2 (20.3 ± 0.9 kHz) than in DE1 (19.6 ± 0.7 Hz).
Courting song The courting song is less intense than the calling song but similarly composed of single and double echemes unevenly arranged; series of one component sometimes repeated during several minutes without a clear phrase structure.
Aggressive song Aggressive interactions between males were not observed despite many attempts in laboratory. This song could be absent from the repertoire of C. enkraussi .







| BPBM |
Bishop Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cardiodactylus enkraussi Otte, 2007
| Robillard, Tony 2009 |
Cardiodactylus enkraussi
| OTTE D. 2007: 353 |
| OTTE D. 2007: 32 |