
Anguilla luzonensis Watanabe, Aoyama, and Tsukamoto, 2009
|
publication ID |
https://doi.org/ 10.12782/specdiv.26.31 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE3543-FFDD-FE60-4927-F152B916FB4F |
|
treatment provided by |
Felipe (2021-11-29 22:07:30, last updated by Plazi 2023-11-08 08:36:24) |
|
scientific name |
Anguilla luzonensis Watanabe, Aoyama, and Tsukamoto, 2009 |
| status |
|
Anguilla luzonensis Watanabe, Aoyama, and Tsukamoto, 2009 View in CoL
[New standard Japanese name: Uguma-unagi] ( Figs 1 View Fig , 2 View Fig ; Tables 1, 2)
Materials examined. Six specimens: the Hiji River,
Okinawa-jima Island , Okinawa Prefecture, 26°43′48″N, 128°10′10″E, KYUM-PI-05433, 48.4 mm TL, 9 October 2018: KYUM-PI-05434, 49.8 mm TL, 30 August 2019 ( Fig. 1 View Fig ); KYUM-PI-05435, 55.1 mm TL, 31 August 2019; KYUM- PI-05436, 50.4 mm TL, 31 August 2019; KYUM-PI-05437, 51.8 mm TL, 29 October 2019; KYUM-PI-05438, 52.4 mm TL, 29 October 2019 GoogleMaps .
Description. Counts and measurements are shown in Table 1. The abdominal and caudal vertebrae could not be counted in all specimens, and the total vertebrae could not be counted in specimens KYUM-PI-05433, 05437, and 05438. Body elongate, slender, subcylindrical in front, laterally compressed in tail ( Fig. 1 View Fig ). Head short, about one eighth of total length, depressed anteriorly. Snout rather short, its tip broadly rounded. Eyes small, circular, located above the edge of upper jaw. Anterior nostril tubular, located near tip of snout. Posterior nostril tubular, located in front of eye. Upper jaw shorter than lower jaw. Lips well developed. Gill opening located in front of pectoral base, crescent in shape, as wide as width of pectoral base. Scales undeveloped. Dorsal, anal and pectoral fin rays well-developed and well-differentiated. Pectoral fins small, their posterior margin rounded. Anal fin base just behind anus. Dorsal fin base located slightly behind center between tip of pectoral fin and anal fin base. Dorsal and anal fins moderately elevated, slightly reducing in height posteriorly, but posteriormost portion expanded, confluent with small caudal fin.
Color when fresh. Body uniformly transparent, a dense belt of melanophores on the caudal peduncle along the mediolateral line, very little pigmentation onto the caudal fin ( Fig. 2 View Fig ); pigment on nerve cord, more developed on the caudal side. Based on Tesch (2003), the ontogenetic stage categorization in the glass eels of six specimens A. luzonensis had melanophore pattern consistent with stage VA for KYUM-PI-05438, stage VB for KYUM-PI-05434–05437, and stage VIB for KYUM-PI-05433.
Color in ethanol. Body uniformly ivory, a large patch of diffused melanophores on the caudal peduncle.
Distribution. Adult A. luzonensis has been recorded only from Luzon Island in the northern Philippines (holotype; Watanabe et al. 2009). Glass eels of this species have been recorded from Luzon Island and Mindanao Island in the Philippines, and Taiwan ( Aoyama et al. 2015; Shinoda et al. 2015; Han et al. 2016; Shirotori et al. 2016).
Habitat and biology. This species was firstly collected from a small stream in the upper reaches of the Pinacanauan River system, a tributary of the Cagayan River on northern Luzon Island, the Philippines (holotype; Watanabe et al. 2009) .
Knowledge on this species is mainly pertains to in its early life history, and we refer to previous studies on the presumed spawning area and transport routes of A. luzonensis . Comparison of otolith increment counts between A. marmorata and A. luzonensis suggested that they may have at least partially overlapping spawning areas ( Han et al. 2016). Previous studies indicate that the estimated spawning area of A. marmorata (12– 17°N, 131– 143°E; Kuroki et al. 2009) overlaps that of A. japonica (12– 16°N, 137– 143°E; Tsukamoto 1992, 2006; Kuroki et al. 2006, 2009). The spawning area of A. luzonensis may also be in these ranges ( Han et al. 2016). In fact, Kuroki et al. (2012) reported that a 29.2 mm leptocephalus of A. luzonensis was collected at 13°N, 140°E in 2009. A simulated tracer experiment indicated that leptocephali of A. luzonensis may reach the North Equatorial Current bifurcation site when the Kuroshio Current is intensified, and leptocephali of A. luzonensis are likely to be transported by the northward-flowing Kuroshio Current in the direction of Luzon Island ( Han et al. 2016). Therefore, A. luzonensis are likely to be transported westward from their spawning areas by the North Equatorial Current, and then transported to Okinawa-jima Island by the Kuroshio Current.
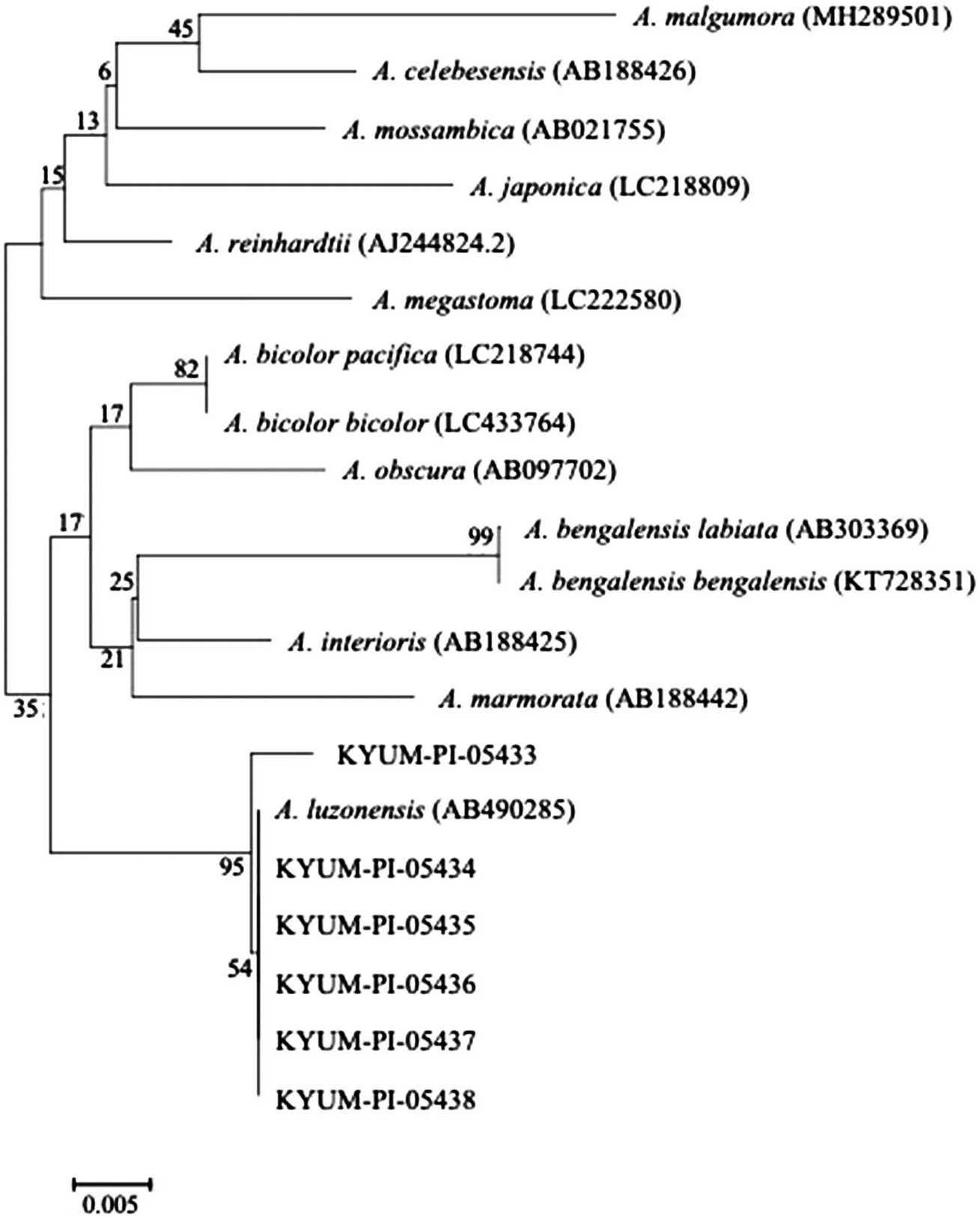
Remarks. The specimens collected as a part of our monthly sampling for ecological studies of glass eels on Okinawa-jima Island, Okinawa Prefecture, the specimens were identified as A. luzonensis by the combination of morphological and molecular methods. Morphometric and caudal pigmentation characteristics of all known species and subspecies of Anguilla in Japan are shown in Table 2 ( Tzeng 1982). The glass eel of A. luzonensis is easily distinguished from A. b. pacifica by ADL% TL value of>5% (vs. <5%), and from A. japonica by the presence of caudal pigmentation (vs. absence). However, the glass eels of A. luzonensis and A. marmorata are not completely distinguished because they have the same caudal pigmentation pattern and some ADL% TL value of overlap ( Leander et al. 2012). According to Leander et al. (2012), among specimens with caudal pigmentation, those with ADL% TL values>13% are A. marmorata, and those with values of <13% are classified as A. marmorata or A. luzonensis . Therefore, specimens with an ADL% TL <13% are distinguished between A. marmorata and A. luzonensis using molecular methods. A neighborjoining tree based on the genetic sequences is shown in Fig. 3 View Fig . As a result of comparison with other species of Anguilla in Indo-Pacific waters based on 16S sequences, the sequences of the six specimens were found to be contained in the clade of A. luzonensis . Accordingly, the six specimens were identified as A. luzonensis based on their morphometric characteristics and DNA sequences. In present study, we followed Leander et al. (2012) to identify A. luzonensis glass eels using ADL% TL value of <13%. However, Shirotori et al. (2016) reported that the range of ADL% TL for A. luzonensis glass eels was 9.6–14.7 (n=4), and included specimen with ADL% TL value of>13%. On the basis of the above report, key to the species of Anguilla (glass eel) in Japan is presented.
In Japan, there have been reports of exotic eel species imported for aquaculture from Europe and Southeast Asia escaping or being released from eel culture ponds into the rivers ( Tabeta et al. 1976, 1977; Arai et al. 2017). Eels are currently cultured in a pond in Kin Town, Okinawa-jima Island. However, there are no eel culture ponds around the Hiji River Basin, where the specimens were collected. Thus, our specimens are not escapees from culture ponds.
In Watanabe (2019), a Japanese name of “Ruson-unagi” was used. However, because this Japanese name is based on unpublished data, it is not possible to determine retrospectively which species is A. luzonensis , making “Ruson-unagi” used by Watanabe (2019) ineligible as a standard Japanese name. Therefore, we herein propose a new standard Japanese name, “Uguma-unagi” for this species based on the specimens (KYUM-PI-05434). The proposed species name is based on the body color of adult of this species [see photo in Wanatabe et al. (2009)] and is a combination of “uguma” meaning “sesame” in the Okinawan dialect to express the irregular mottled coloration pattern on the body of the species, and “unagi” meaning “eel” in Japanese.
Aoyama, J., Yoshinaga, T., Shinoda, A., Shirotori, F., Yambot, A. V., and Han, Y. S. 2015. Seasonal changes in species composition of glass eels of the genus Anguilla (Teleostei: Anguillidae) recruiting to the Cagayan River, Luzon Island, the Philippines. Pacific Science 69: 263 - 270.
Arai, K., Itakura, H., Yoneta, A., Yoshinaga, T., Shirotori, F., Kaifu, K., and Kimura, S. 2017. Discovering the dominance of the non-native European eel in the upper reaches of the Tone River system, Japan. Fisheries Science 83: 735 - 742.
Han, Y. S., Lin, Y. F., Wu, C. R., Iizuka, Y., Castillo, T. R., Yambot, I. U., Mamalangkap, M. D., and Yambot, A. V. 2016. Biogeographic distribution of the eel Anguilla luzonensis: dependence upon larval duration and oceanic currents. Marine Ecology Progress Series 551: 227 - 238.
Kuroki, M., Aoyama, J., Miller, M. J., Wouthuyzen, S., Arai, T., and Tsukamoto, K. 2006. Contrasting patterns of growth and migration of tropical anguillid leptocephali in the western Pacific and Indonesian Seas. Marine Ecology Progress Series 309: 233 - 246.
Kuroki, M., Aoyama, J., Miller, M. J., Yoshinaga, T., Shinoda, A., Hagihara, S., and Tsukamoto, K. 2009. Sympatric spawning of Anguilla marmorata and Anguilla japonica in the western North Pacific Ocean. Journal of Fish Biology 74: 1853 - 1865.
Kuroki, M., Miller, M. J., Aoyama, J., Watanabe, S., Yoshinaga, T., and Tsukamoto, K. 2012. Evidence of offshore spawning for the newly discovered anguillid species Anguilla luzonensis (Teleostei: Anguillidae) in the western North Pacific. Pacific Science 66: 497 - 508.
Leander, N. J., Shen, K. N., Chen, R. T., and Tzeng, W. N. 2012. Species composition and seasonal occurrence of recruiting glass eels (Anguilla spp.) in the Hsiukuluan River, eastern Taiwan. Zoological Studies 51: 59 - 71.
Shinoda, A., Yoshinaga, T., Aoyama, J., Tsuchida, G., Nakazato, S., Ishikawa, M., Matsugamoto, Y., Watanabe, S., Azanza, R. V., and Tsukamoto, K. 2015. Early life history of the Luzon mottled eel Anguilla luzonensis recruited to the Cagayan River, Luzon Island, the Philippines. Coastal Marine Science 38: 21 - 26.
Shirotori, F., Ishikawa, T., Tanaka, C., Aoyama, J., Shinoda, A., Yambot, A. V., and Yoshinaga, T. 2016. Species composition of anguillid glass eels recruited at southern Mindanao Island, the Philippines. Fisheries Science 82: 915 - 922.
Tabeta, O., Takai, T., and Matsui, I. 1976. Record of short finned eel from Nagata River, Shimonoseki, Japan. Bulletin of the Japanese Society of Scientific Fisheries 42: 1333 - 1338.
Tabeta, O., Takai, T., and Matsui, I. 1977. [Elvers imported into Japan for eel culture] The Aquiculture 24: 116 - 122. [In Japanese]
Tesch, F. W. 2003. Pigmentation. Pp. 13 - 17. In: Tesch, F. W. (Ed.) The Eel. Blackwell Science, London.
Tsukamoto, K. 1992. Discovery of the spawning area for Japanese eel. Nature 356: 789 - 791.
Tzeng, W. N. 1982. Newly record of the elver, Anguilla celebesensis Kaup, from Taiwan. Chinese Bioscience 19: 57 - 66. [In Chinese with English abstract]
Watanabe, S., Aoyama, J., and Tsukamoto, K. 2009. A new species of freshwater eel Anguilla luzonensis (Teleostei: Anguillidae) from Luzon Island of the Philippines. Fisheries Science 75: 387 - 392.
Watanabe, S. 2019. [The ecology of eels]. Pp. 17 - 48. In: Tsukamoto, K. (Ed.) Unagi no Kagaku [The Science of Eels]. Asakura Publishing Co., Ltd., Tokyo. [In Japanese]

Fig. 1. Fresh specimen of Anguilla luzonensis glass eel, KYUM-PI-05434, 49.8mm in total length, collected from Okinawa-jima island, Japan.

Fig. 2. Caudal fin and caudal pigmentation pattern of Anguilla luzonensis glass eel (fresh specimens), KYUM-PI-05434.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |