
Oligosita balcluthae Viggiani et Laudonia
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4039.4.8 |
|
publication LSID |
lsid:zoobank.org:pub:D86C0159-4D25-4C45-BB99-4E043E0E6D72 |
|
DOI |
https://doi.org/10.5281/zenodo.6104286 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE879C-FFB2-FFFC-FF3B-FA3AFD89FD18 |
|
treatment provided by |
Plazi |
|
scientific name |
Oligosita balcluthae Viggiani et Laudonia |
| status |
sp. nov. |
Oligosita balcluthae Viggiani et Laudonia n. sp.
Figs 1 View FIGURE 1. a – f , 2 View FIGURE 2. a – c
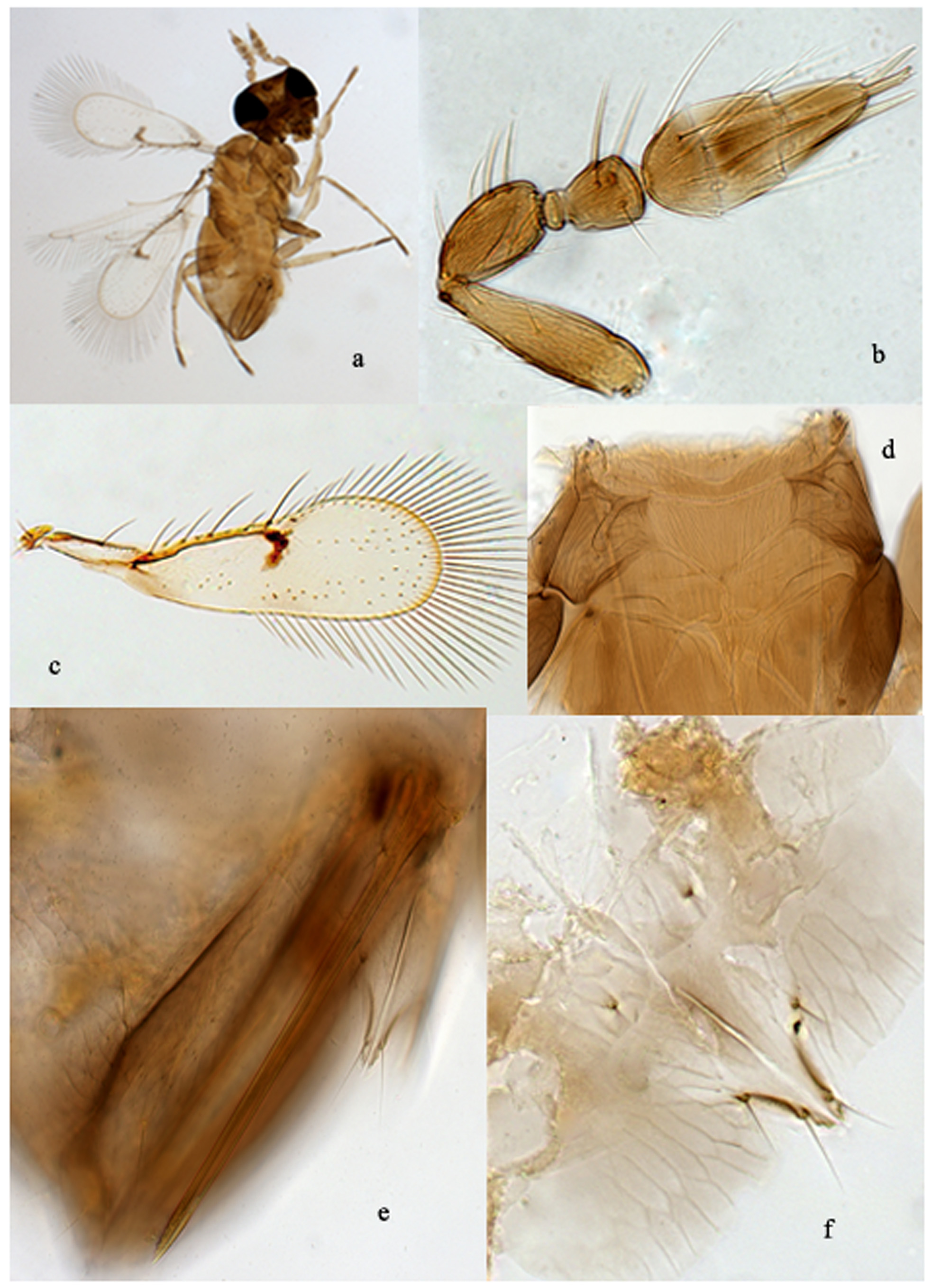
Description. Female ( Fig. 1a View FIGURE 1. a – f ). Body length: 0.65 mm (SD: 0.060; n: 20). Body honey yellow, ocelli and eyes black, head below the level of lower margin of eyes brown, some infuscation may be present laterally and ventrally on prothorax, and laterally on mesosoma and metasoma. Ovipositor sheaths brown. Fore wing ( Fig. 1c View FIGURE 1. a – f ) blade lightly infuscate from base to distal margin; additional black spots present at level of premarginal vein and stigmal vein. Legs with coxae and last tarsomeres brown.
Ratio of head: mesosoma: metasoma length = 5:8:14. Head slightly wider than mesosoma, about twice as wide as long. Mandibles each with 2 teeth and an internal truncation. Maxillary palpus one segmented, twice as long as wide, with a stout process of the same length and a terminal seta twice as long as palpus. Labial palpus vestigial, with a long seta as long as terminal seta on maxillary palpus. Antenna ( Fig. 1b View FIGURE 1. a – f ) inserted just above level of lower ocular line; scape rather narrow, as long as club; pedicel slightly wider than scape; anellus transverse, very small; funicle segment narrowest basally, narrower and shorter than pedicel (10:17); club widest at level of distal part of first segment and base of second segment, the latter slightly longer than the third; apical process a little shorter than third claval segment (8: 10). Setae and sensilla of antenna as in Fig. 1b View FIGURE 1. a – f .
Pronotum short, as long as metanotum, each lateral angle with a seta. Mid lobe of mesoscutum a little wider than long (40:35), with shallow, longitudinal, net-like sculpture and with one seta half as long as scutellum at midlevel near lateral margin; scutellum one-third length of mid lobe of mesoscutum, with a pair of setae and two pores, each pore placed internally near base of seta and distant from each other by distance equal to length of seta. Axilla with one seta. Metanotum ( Fig. 1d View FIGURE 1. a – f ) very short, about one-third as long as scutellum, with some striation. Propodeum ( Fig. 1d View FIGURE 1. a – f ) as long as scutellum, with a triangular mid lobe and longitudinal striation also extending to lateral parts of the sclerite with reticulate sculpture; spiracle situated a little more than own diameter from anterior margin of propodeum. Fore wing ( Fig. 1c View FIGURE 1. a – f ) 3.4× as long as broad; venation extending about 3/5 wing length; ratio of submarginal: premarginal: marginal: stigmal veins = 18:8:20:6. Costal cell narrow, twice as long as marginal vein, with one seta on anterior margin at level of mid-length of premarginal vein; veins with rather long setae, one seta on submarginal vein about as long as premarginal vein, one at base of premarginal vein about as long as seta on submarginal vein, and three on marginal vein, the distal one the longest. Fore wing blade with distal margin not regularly rounded, with a few (4–6) microsetae along posterior margin opposite of marginal vein; disc distally beyond level of stigmal vein with a row of submarginal setae, with 2 to 3 irregular rows of setae extending from uncus to distal margin, and another 2 to 3 irregular rows extending just above posterior margin of disc from level of stigmal vein to distal margin; fringe well developed; maximum length of cilia along anal margin about as long as maximum width of disc. Hind wing with a row of microsetae just behind anterior margin. Front leg with tarsus a little longer than tibia (25:20), tarsomeres subequal in length and width, with basitarsomere shorter than last tarsomere (6:8); middle leg with basitarsomere longer than distal tarsomere (10:8); hind leg with tarsomeres as for middle leg and tibial spur about half as long as basitarsomere. Metasoma ovate ( Fig. 1a View FIGURE 1. a – f ), as long as head and mesosoma combined; urotergites with shallow longitudinal reticulation. Ovipositor with base inserted at level of the fourth urite, not extruded; third valvulae very short, only 0.13 total ovipositor length; ratio of ovipositor: front tibia length = 4:2; hypogynium distal margin reaching basal half of ovipositor ( Fig. 1e View FIGURE 1. a – f ) and with ventral surface as in Fig. 1f View FIGURE 1. a – f .
Male. Similar to female in colour and morphological features. Body length 0.56 mm (DS: 0.072; n: 20) a little shorter than that of female. Metasoma distally less pointed and with some dusky spots. Antenna ( Fig. 2a View FIGURE 2. a – c ) without apical process. Male genitalia inserted at level of VIII urite; tubular, in lateral view curved ventrally ( Fig. 2b View FIGURE 2. a – c ), the basal half a little wider than the last distal part; aedeagal apodemes ( Fig. 2c View FIGURE 2. a – c ) short, as long as width of aedeagus body. Dimensions of the male genitalia: total length on average 0.10 mm (SD: 0.003 mm; n: 15); basal width: 0.01 mm (SD: 0.001 mm; n: 15); apodeme length: 0.01 (SD: 0.001 mm; n: 15).
Etymology. The name is in reference to the host genus Balclutha .
Material examined. Holotype (♀), on slide, Catania, 11.VII.2014, Piazza Michelangelo (37°31'22'' N, 15°05'34'' E); 43 m a.s.l., from egg of Balclutha brevis , leg. S. Bella. Allotype (♂): Catania, 18.X.2014, same locality as holotype.
Paratypes: 5 ♀, same data as holotype; 3 ♀ and 7 ♂ on slide, same data as allotype; 1 ♀ and 1 ♂ on card, from parasitized eggs collected in Catania, III.2015, emerged in the Department of Agriculture-BIPAF laboratory on 8.IV.2015.
Additional material: 27 ♀ and 8 ♂ in alcohol, same data as holotype; 40 ♀ and 8 ♂, dried specimens, same data as holotype deposited in the entomological collection of the Department of Agriculture-BIPAF University of Naples “Federico II”, Portici, Napoli, Italia. 43 ♀ in alcohol and 13♀ dried specimens, 28 ♂ in alcohol and 8 ♂ dried specimens are preserved in the laboratory of CREA-ACM Acireale (Catania). 183 ♀ and 97 ♂ in alcohol are deposited in the entomological collection of the Department of Biological, Geological and Environmental Sciences, Section of Animal Biology "Marcello La Greca", University of Catania.
The holotype, 5 ♀ and 4 ♂ paratypes and all additional material are deposited in the entomological collection of the Department of Agriculture-Division of Biology and Protection of Agricultural and Forest Systems ( BIPAF) University of Naples “Federico II”, Portici, Napoli, Italia; 1♀ and 1 ♂ paratype in the National History Museum, London, England, U.K.; 1 ♀ and 1 ♂ paratype in the University of California, Riverside, California, USA; 1 ♀ and 1 ♂ paratype in the National Museum of Natural History, Washington, D. C.
Taxonomic notes. This new species belongs to the collina group (Viggiani 1976) of Oligosita Walker as redefined by Pinto and Viggiani (2004). Among known species belonging to this group, which is characterized mostly by antenna and fore wing shape, O. balcluthae appears very similar to O. biscrensis Nowicki (1935) . The latter species is known only from its original description, based on a female collected in a palm orchard of Biscra (Northern Sahara), Algeria, June 24, 1931. The type of O. biscrensis is preserved in the entomological collection of the Dipartimento di Entomologia e Zoologia agraria “Filippo Silvestri” ( DEZA), Portici (NA), Università degli Studi di Napoli “Federico II”. It was dissected and originally mounted on three cards; each with a circular hole covered with glass on both sides, but was remounted by Viggiani on two slides using balsam-phenol as the medium. Oligosita biscrensis should be added to the list of species in the S. Nowicki collection preserved in the above mentioned institution by Viggiani (2011).
Specimens of O. balcluthae can be distinguished from O. biscrensis by the lower part of the head being dark, the fore wing fringe being about as long as the maximum discal width, and the ovipositor being only twice as long as the front tibia. The holotype of O. biscrensis has the head completely yellow, the longest setae of the fore wing fringe shorter than the maximum discal width (2/3), and the ovipositor three times as long as the front tibia ( Fig. 2 View FIGURE 2. a – c d).
Male genitalia of O. balcluthae show the typical shape known for other species of the collina group ( Viggiani 1971).
Molecular analyses. The 28S-D2 and ITS 2 regions were successfully amplified and sequenced. ITS 2 was used to provide molecular analysis of the new species. rDNA genes or nucleotide regions have proven to be useful in taxonomic studies for various taxa and one of the most important target regions used in molecular analysis was ribosomal DNA which is relatively less changed and is composed of 28S D2 gene regions and internal spaces ITS 2. The sequence analysis of ITS 2 rDNA has been described in several studies as a tool for Trichogrammatidae identification as well (Kan et al. 1996; Pinto et al. 1997, 2002; Sayed et al. 2011; Sumer et al. 2011). 28S rDNA coding rRNA is accepted as a highly conserved gene region during evolutionary processes, so it is used to construct phylogenetic trees especially for higher categories. Ribosomal ITS 2 is a non-coding region located between 5.8S and 28S ribosomal DNA regions. Because this region is thought to change rapidly, it has been used commonly in population genetics, separation of cryptic species, and similar species. For Oligosita , PCR product sizes of the region ITS 2 varied from 460 to 510 bp ( Ávila-Rodríguez et al. 2013).
The 28S-D2 sequences (published in gene bank, accession number: BankIt1831844 Seq1 KT124387 View Materials ) correspond completely with the 28S-D2 sequences published by Gillespie et al. (2005) for Oligosita .
GRAPH 1. Number of adults of O. balcluthae (N) emerged from a sample of 10 P. setaceum ears collected in two locations of Catania (Piazza Michelangelo and Via Giovannino).
ITS 2 sequences did not show polymorphic sites, but all specimens had a common origin, collected in the same place and from the same leafhopper eggs. In fact, from the multiple sequence alignment only one sequence was obtained which contained 498 pb (sequence published in gene bank, accession number: BankIt1831844 Seq2 KT124388 View Materials ), and different from the observed data for Oligosita sp1 and Oligosita sp 2 in Ávila-Rodríguez et al. (2013). It was not possible to compare the ITS 2 sequences to other sequences obtained from Oligosita due to lack of published data.
Biology. Oligosita balcluthae oviposits in the eggs of the host, which measure 0.7–0.8 mm in length and 0.2– 0.3 in width, located on the internal side of the glumes in groups of 2 or 3 elements. A single parasitoid develops in each host egg ( Fig. 2 View FIGURE 2. a – c e). The parasitization starts to be evident during the last larval stage when the host egg chorion becomes grey or brown ( Fig. 2 View FIGURE 2. a – c f). The pupal stage, with black eyes, is clearly visible. The healthy leafhopper eggs during embryonic development show red eye spots. Ears of P. setaceum sampled from June 6, 2012 to May 28, 2013, showed that O. balcluthae adults emerged almost all year around, except during winter, when the parasitoid develops slowly its young stages. In 2015 a sample of P. setaceum ears collected in the second half of March showed adult O. balcluthae emerged at the beginning of April. The first annual emergence of the parasitoid matches the presence of fresh eggs of the leafhopper host. The abundance of O. balcluthae emerging from the P. setaceum ears is shown in the Graph 1.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.