
Rhynchozoon maculosum, Dick & Grischenko, 2016
|
publication ID |
https://doi.org/10.1080/00222933.2016.1253797 |
|
DOI |
https://doi.org/10.5281/zenodo.4333644 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE87C2-D108-5C19-63D7-FF74FE8DFC40 |
|
treatment provided by |
Carolina |
|
scientific name |
Rhynchozoon maculosum |
| status |
sp. nov. |
Rhynchozoon maculosum sp. nov.
( Figures 35 View Figure 35 and 36 View Figure 36 (a – d))
Etymology
The specific name is an adjective (Latin maculosus: mottled, full of spots), referring to the many areolae speckling the colony, giving it a ‘ peppered ’ appearance.
Material examined
Holotype: NSMT-Te 1182, dried specimen, SES site; NSMT-Te 1183 ( SES-14 ), part of same colony as NSMT-Te 1182, bleached, on SEM stub . Paratypes: NSMT-Te 1184 ( SES- 15),
bleached, on SEM stub; NSMT-Te 1185 ( SES-18 ), bleached, on SEM stub; NSMT-Te 1186 ( SES-19 ), bleached, on SEM stub; NSMT-Te 1187 ( SES-51 ), bleached, on SEM stub; NSMT- Te 1188 ( MIN-20-1 ), bleached, on SEM stub; NSMT-Te 1189, paratype lot, four dried specimens, SES site; NHMUK 2016.5 About NHMUK . 13.73 ( MIN-20 ), bleached, on SEM stub; NHMUK 2016.5.13.74-77, four dried specimens, SES site . Other material: NSMT-Te 1190 ( SES-47 ), bleached, on SEM stub with Rhynchozoon scimitar ; NSMT-Te 1191, 32 dried specimens, SES site; NSMT-Te 1192, 14 dried specimens, REEF site; NSMT-Te 1193, 12 dried specimens, MIN site; NSMT-Te 1194, dried specimen with externally calcified epibiont, SES site .
Measurements
AzL, 0.46 – 0.63 (0.543 ± 0.048); AzW, 0.27 – 0.36 (0.327 ± 0.026) (n = 18, 4). OrL, 0.092 – 0.128 (0.106 ± 0.009); OrW, 0.097 – 0.120 (0.106 ± 0.006) (n = 18, 4). OvL, 0.14 – 0.25 (0.183 ± 0.028); OvW, 0.17 – 0.26 (0.224 ± 0.021) (n = 15, 2). Largest colony observed 40 × 35 mm.
Description
Colony ( Figure 35 View Figure 35 (a)) forming a unilaminar, encrusting sheet; white or off- white (greyish or faintly tinged with orange or yellow); embryos light orange in dried specimens. Zooids at margin ( Figure 35 View Figure 35 (b, c)) irregularly hexagonal; distinct, delineated by a shallow groove and a suture line flanked by areolae; even with age, zooidal outlines remain more or less discernable. Frontal wall without pseudopores and with four to six circular or oval areolae along each lateral margin. In marginal zooids, frontal wall convex, smooth, somewhat uneven. With increasing secondary calcification, frontal wall rapidly thickens, remaining smooth in surface texture but becoming more rugose, with scattered low, rounded tubercles or nodules; areolar openings increase in size. Primary orifice soon becomes immersed as zooids age, with an asymmetrical pseudosinus forming in rim of secondary orifice, to one side or other of midline. Primary orifice (including sinus) is as long as broad, or occasionally slightly broader than long; anter exclusive of sinus is broader than long. Sinus is moderately deep, rounded-Vshaped, extending between small, rounded condyles. Margin of anter is beaded with 14 or 15 (mode, 15; n = 5, 3) small denticles. Oral spines lacking. With age, rim of secondary orifice usually retains pseudosinus and bears from four to eight small, blunt processes ( Figure 35 View Figure 35 (d – f)) that are irregular, conical or cylindrical in shape. Suboral and frontal avicularia occur. Suboral avicularium develops from hemispherical chamber ( Figure 35 View Figure 35 (b)) occupying one-fifth to one-third the frontal area proximal to the orifice. Avicularian rostrum is offset to one side or other of midline ( Figure 35 View Figure 35 (c)), tilted laterally towards orifice, and directed distolaterally; end hooked; crossbar complete; mandible long-triangular. Base of rostrum is extended as a rounded process (a socalled ‘ uncinate process ’) flanking the orificial pseudosinus and jutting into the peristome. With increased secondary calcification, suboral avicularium lies at edge of secondary orifice, or entirely within peristome. Frontal avicularia ( Figure 35 View Figure 35 (e, f)) roughly same size as suboral avicularia; one to four per zooid, or lacking. Rostrum slightly raised from frontal plane distally, not hooked; crossbar complete; mandible acute, about twice as long as wide, directed laterally, distolaterally, proximolaterally or occasionally medially. Rostrum of frontal avicularia is occasionally diamond-shaped, but usually rounded at proximal end. Ovicell initially subimmersed ( Figure 35 View Figure 35 (e)) but later becoming endozooidal ( Figure 35 View Figure 35 (f)). Ooecium broader than long; proximal face with transversely long-elliptical, semicircular or oval zone of membranous (non-calcified) ectooecium that, in cleaned specimens, becomes a window ( Figure 35 View Figure 35 (e), arrowhead) exposing the entooecium; labellum ( Figure 35 View Figure 35 (e), arrow) narrow, usually complete. Ancestrula not observed.
Remarks
This species is similar to Rhynchozoon rostratum ( Busk 1856) , originally described from the eastern Pacific at Mazatlán, Mexico. The two species share the following characters: the suboral and frontal avicularia are similar in size and shape; the suboral avicularia are often immersed within the peristome, have an ‘ uncinate process ’ at the base of the rostrum, and have a hooked tip; boundaries between mature zooids are indicated by marginal pores; the secondary orifice bears a deep pseudosinus and is surrounded by conspicuous peristomial processes; and oral spines are lacking ( Hastings 1930; Osburn 1952). Rhynchozoon rostratum differs from R. maculosum in having the exposed entooecial area on the proximal face of the ooecium large, semicircular and opaque white, with the labellum typically lacking or incomplete. In R. maculosum , the entooecial zone is markedly transversely long-elliptical or oval, and not conspicuously white, and the labellum is usually complete. Rhynchozoon rostratum further differs from R. maculosum in having a shallower orificial sinus, a shorter mandible on the suboral avicularium, and fewer processes around the peristome. Finally, Hastings (1930) reported colonies of R. rostratum in Panama to have a faintly greenish ectocyst, whereas our dried specimens are faintly yellowish, although this may not be a useful diagnostic character ( Soule and Soule 1964). It is noteworthy that an opaque-white zone of exposed entooecium on the ovicell is not in itself diagnostic of R. rostratum , as it also occurs in R. scimitar sp. nov. described below.
Hastings (1930) appears to have been the only author subsequent to the original description to examine Busk ’ s type specimens of R. rostratum from Mazatlán, and her description of conspecific material from Panama perhaps warrants more weight than other descriptions, among which there are discrepancies. According to Osburn (1952), R. rostratum has a low-boreal to tropical distribution in the eastern Pacific, occurring from Point Conception, California, southward to the Galapagos. He considered R. rostratum also to occur in the western Atlantic under the junior synonym R. verruculatum ( Smitt 1873) , although Hastings (1930), Winston (1984), and Winston and Heimberg (1986) have established that these are different species. Some of the records of R. rostratum in the eastern Pacific are questionable. Soule and Soule (1964) mentioned marginal zooids with two to four oral spines in material they identified as R. rostratum from Scammon ’ s Lagoon, Baja California, Mexico. Soule et al. (1995) reported R. rostratum from the Santa Barbara Channel, southern California, USA, to have two oral spines on marginal zooids and a circular or transversely oval entooecial area on the ovicell, with a complete labellum, characters that conflict with Hastings ’ s (1930) redescription of R. rostratum from Panamanian material. Specimens from Kodiak, Alaska, reported as R. rostratum ( Dick and Ross 1988) are instead R. tumulosum ( Hincks 1882b) (Dick et al. 2005; Dick and Mawatari 2005). We suspect that some of the inconsistencies (e.g. spines present or absent; variation in colony colour and the depth of the oral sinus) among descriptions of nominal R. rostratum in the eastern Pacific actually reflect interspecific differences, and that SEM examination of a broad range of material will resolve nominal R. rostratum into a number of morphologically distinct species.
Indo-West Pacific records of R. rostratum have now been referred to other species. Tilbrook et al. (2001) considered R. rostratum reported from Java ( Winston and Heimberg 1986) and the Philippines (Scholz 1991) actually to be R. splendens Hayward, 1988 , and Gordon (2009) erected a new species, R. zealandicum , for New Zealand material previously reported ( Gordon 1970) as R. rostratum .
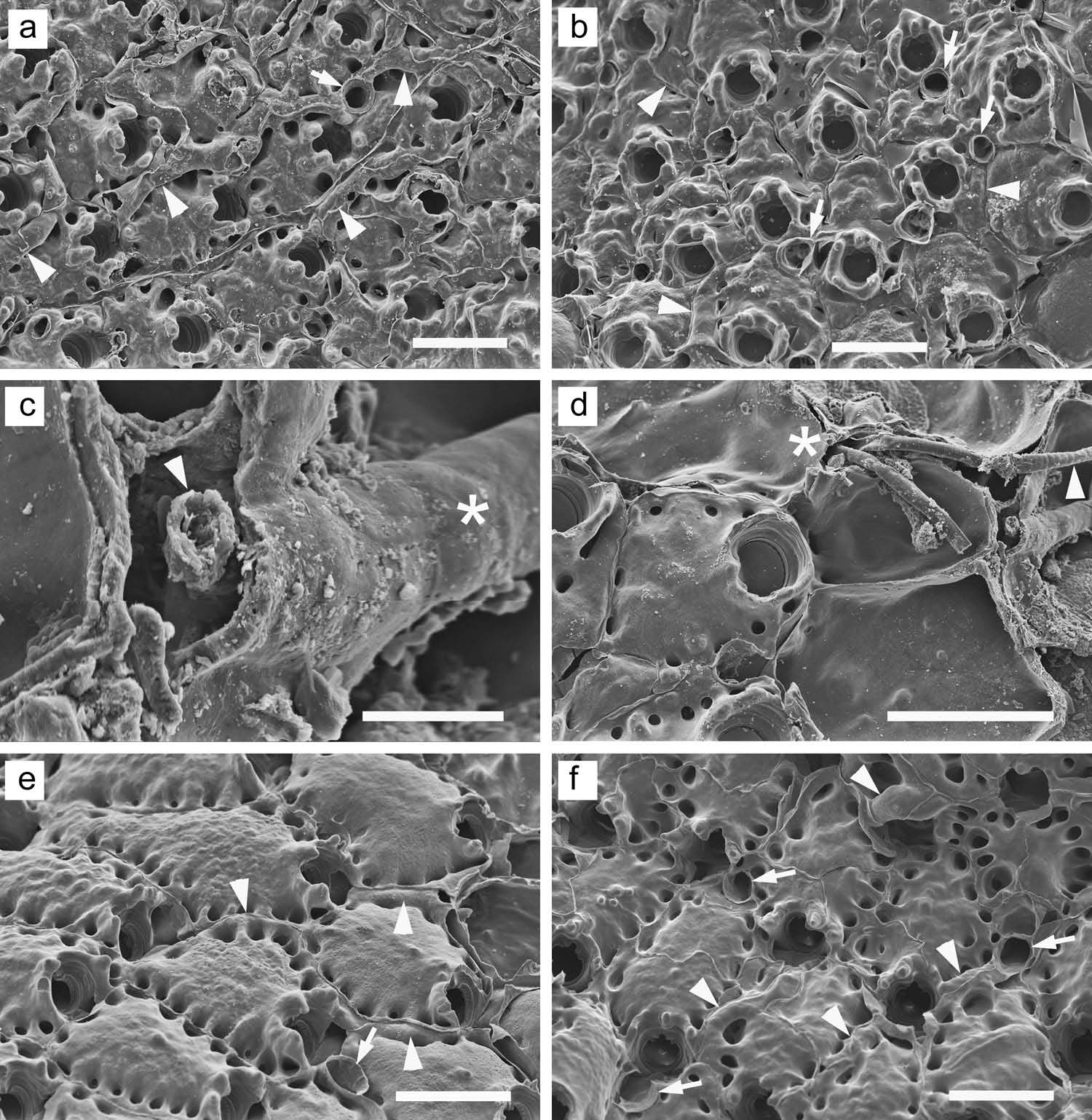
Two of the six colonies of R. maculosum we examined by SEM had a filamentous, ramifying epibiont in the interzooidal grooves ( Figure 36 View Figure 36 (a – d)). The bryozoan zooids deposited a thin layer of gymnocystal calcification beneath the filaments of the epibiont growing on the colony surface, encased them completely in calcified tubes, or caused the tubes to become completely immersed between zooids by depositing secondary calcification over them (arrowheads, Figure 36 View Figure 36 (a, b)); in the last case, the tubes emerged to the surface by way of irregularly occurring circular openings (arrows, Figure 36 View Figure 36 (a, b)). Examination of an unbleached specimen showed that the soft filaments of the epibiont are hollow (arrowhead, Figure 36 View Figure 36 (c)), and one emergent filament produced an annulated section reminiscent of a hydroid stalk (arrowhead, Figure 36 View Figure 36 (d)), leading us to speculate that the ramifying epibiotic filaments were hydroid stolons. The epibiont affected the bryozoan colonies in at least two ways: the extensive surficial gymnocystal calcification produced to ‘ wall off ’ the epibiont must have incurred an energetic cost, and the epibiont appears to have inhibited the production of avicularia. In one affected colony, zooids produced no avicularia at all; in the other, while some zooids produced avicularia, both the suboral and frontal types were reduced in frequency.
Occurrence
This species was common or abundant at all three sites ( Table 1), the only known localities.

| SES |
Southeastern Shanxi Teachers School |
| SES- |
Southeastern Shanxi Teachers School |
| NHMUK |
Natural History Museum, London |
| MIN |
University of Minnesota |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Inovicellina |
|
SuperFamily |
Buguloidea |
|
Family |
|
|
Genus |