
Celleporaria triangula Seo, 1994
|
publication ID |
https://doi.org/ 10.1080/00222933.2016.1253797 |
|
DOI |
https://doi.org/10.5281/zenodo.4333641 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE87C2-D142-5C43-63DE-FCCDFC9EFA73 |
|
treatment provided by |
Carolina |
|
scientific name |
Celleporaria triangula Seo, 1994 |
| status |
|
Celleporaria triangula Seo, 1994
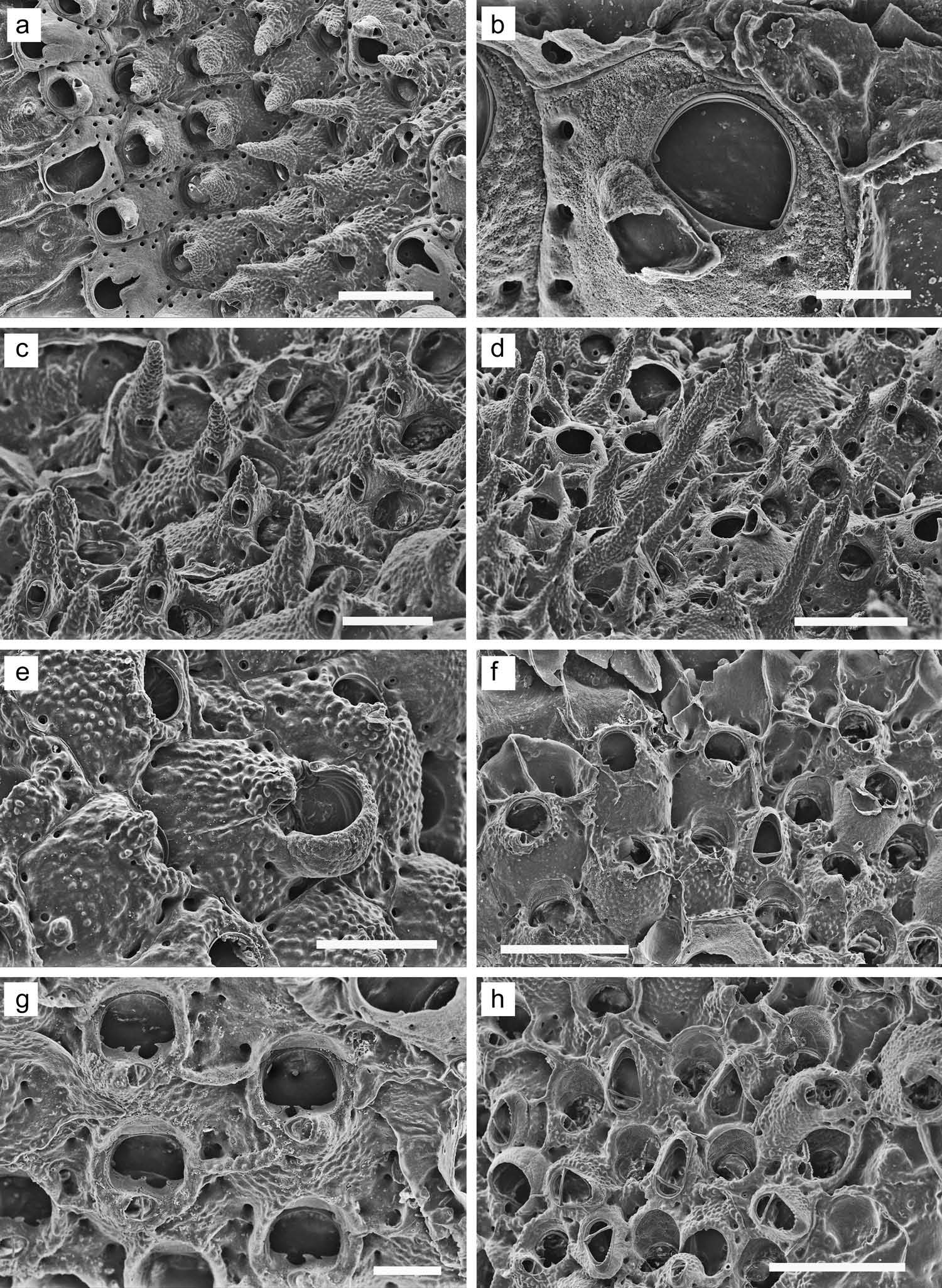
( Figure 14 View Figure 14 (f – h))
Celleporaria triangula Seo, 1994, p. 189 , pls. 1, 2.
Celleporaria triangula: Seo 2005, p. 397 , pls. 120B, 121.
Measurements
AzL, 0.44 – 0.47 (0.455 ± 0.017); AzW, 0.32 – 0.36 (0.338 ± 0.018) (n = 3, 1). OrL (including sinus), 0.12 – 0.16 (0.134 ± 0.014); OrW, 0.14 – 0.18 (0.158 ± 0.012) (n = 13, 1). OvL, 0.21 – 0.24 (0.225 ± 0.016); OvW, 0.25 – 0.26 (0.256 ± 0.002) (n = 3, 1). Largest colony observed 9 mm across.
Material examined
NSMT-Te 1097-a ( SES- 13), bleached, on SEM stub (with Parasmittina soulesi ); NSMT-Te 1098, 15 dried specimens, SES site; NHMUK 2016.5.13.26-38, 13 dried specimens, SES site.
Description
Colony encrusting, multilaminar, forming a low mound. Zooids at colony margin ( Figure 14 View Figure 14 (f)) irregularly hexagonal; zooidal boundaries indistinct in colony interior. Frontal wall coarsely granulated, with three or four small, circular areolae along each lateral margin. Newly budded zooids at margin or interior of colony margin occasionally show one or two spine scars or small, cylindrical spine bases distal to the orifice; most zooids show no trace of spines. Primary orifice ( Figure 14 View Figure 14 (g)) broader than long, roughly D-shaped; with small, blunt, condyles near corners. Proximal orificial margin slightly concave, with three small, falciform or denticulate processes delineating asymmetrical proximal sinuses (one larger, one smaller). Peristome a thick, low rim proximal and lateral to orifice; primary orifice not deeply sunken and usually visible. Small suboral avicularium on raised, sometimes slightly umbonate chamber proximal to orifice, contributing to proximal peristome; rostrum oval, acute to frontal plane, bluntly denticulate around distal margin; crossbar complete; mandible D-shaped, directed frontolaterally. Large interzooidal avicularia are scattered abundantly among autozooids; chamber raised, granulated like frontal wall except at rostral end; rostrum raised distally, bluntly serrate around distal margin; crossbar complete; mandible blunt-triangular. Ovicells ( Figure 14 View Figure 14 (h)) incompletely developed in our material; the most complete is cowlshaped, not yet covered with granulated secondary layer.
Remarks
In her remarks accompanying the original description of C. triangula, Seo (1994) listed a number of Celleporaria species that have denticles on the proximal orificial margin. Taxonomists giving weight to the presence of these denticles have likely misidentified specimens of a variety of species as C. tridenticulata ( Busk, 1881) , C. vagans ( Busk, 1881) or C. aperta ( Hincks, 1882a) , which have been perceived as widely distributed; see Tilbrook (2006) for a discussion of the problem. Tilbrook (2006, pl. 28A, B) illustrated a syntype specimen for C. tridenticulata that has orificial denticles quite different from those in C. triangula , and has lanceolate interzooidal avicularia. Tilbrook (2006, pl. 28C, D) also illustrated a paratype specimen of C. vagans ; the denticles again differ from in C. triangula , and the interzooidal avicularia are spatulate, with a palmate mandible. Harmer (1957) considered C. aperta to have a very broad Indo- Pacific distribution, but both Hincks ’ s (1882a) original description and redescriptions by Winston and Heimberg (1986) and Ryland and Hayward (1992) show the orifice with a small, rounded median sinus lacking a median denticle. The variation that Harmer (1957, p. 674, text-fig. 56) illustrated in the orificial denticles of C. aperta includes some of that seen in C. triangula , and indeed some of the specimens he included in the former may actually be the latter. Celleporaria triangula differs from congeners in the following combination of characters: one or two ephemeral oral spines; orificial denticles delineating major and minor sinuses, with the denticles themselves quite variable; interzooidal avicularia having a blunt-triangular (rather than spatulate, lanceolate or acute) mandible; and a bluntly denticulate distal margin of the rostrum in both the suboral and interzooidal avicularia.
Occurrence
Celleporaria triangula was restricted to the SES site, where it was abundant ( Table 1). This species was previously known only from the vicinity of Cheju Island in the East China Sea, and from the Strait of Korea off the coast of South Korea ( Seo 1994).
Superfamily SMITTINOIDEA Levinsen, 1909 Family SMITTINIDAE Levinsen, 1909
Genus Parasmittina Osburn, 1952
Parasmittina serrula Soule and Soule, 1973
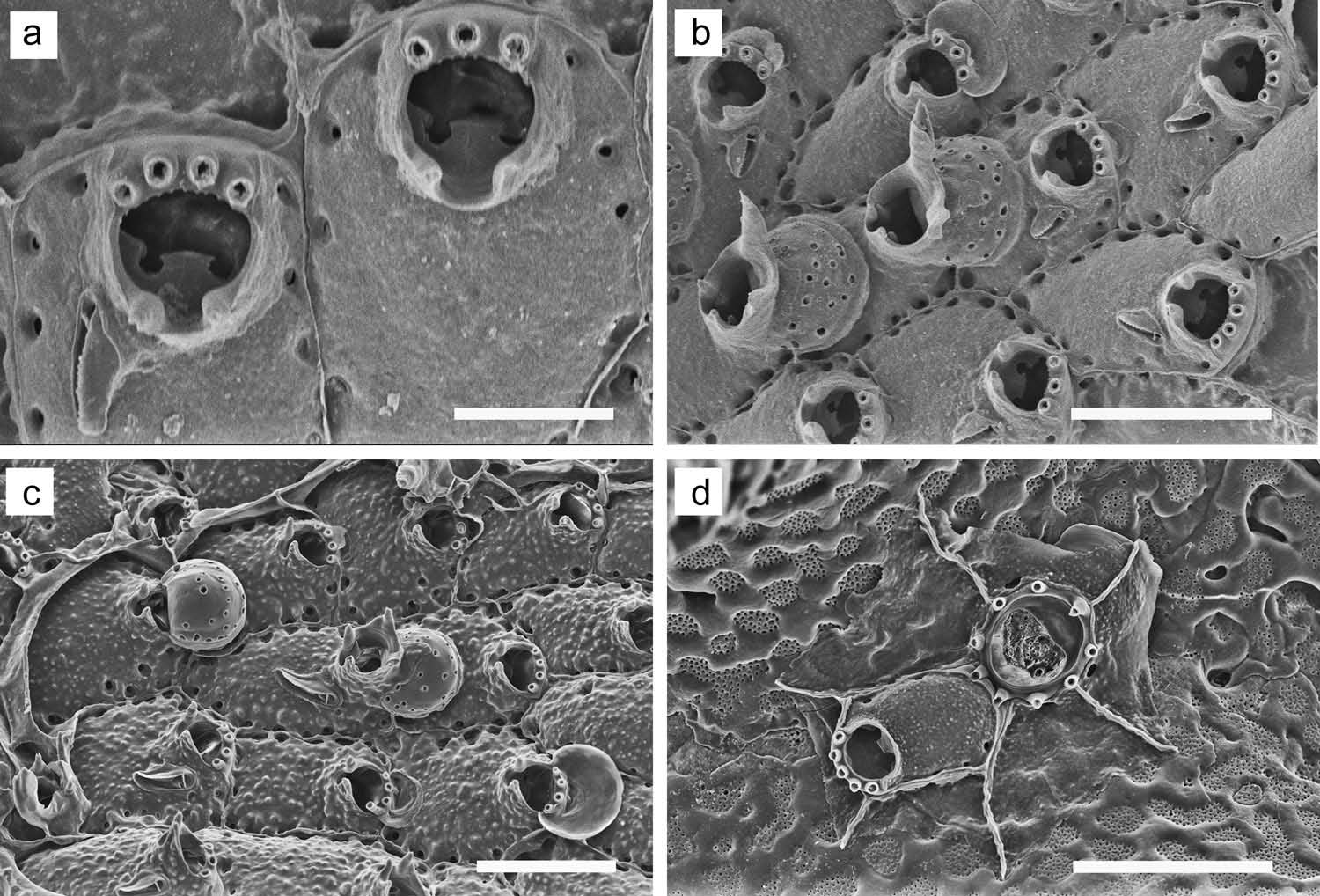
( Figure 15 View Figure 15 )
Parasmittina serrula Soule and Soule, 1973, p. 396 , fig. 3(d – f). Parasmitttina serrula: Gordon 1984, p. 96 , pl. 35B, C. Winston 1984, p. 23, fig. 45. Ryland and Hayward 1992, p. 272, figs 23(e, f) and 24(a). Dick et al. 2006, p. 2221, fig. 9(a – d).
? Parasmittina serrula: Scholz 1991, p. 325 , pl. 29, figs 1, 2 and 4. Tilbrook et al. 2001, p. 76, fig. 14(a, b). Tilbrook 2006, p. 154, pls. 29C, 31A – C.
Material examined
NSMT-Te 1099 ( MIN- 22), bleached, on SEM stub; NSMT-Te 1100 ( MIN- 13), bleached, on SEM stub (with Crepidacantha poissonii ); NSMT-Te 1101, dried specimen, SES site (with Calyptotheca reniformis ).
Measurements
AzL, 0.28 – 0.47 (0.374 ± 0.053); AzW, 0.19 – 0.29 (0.239 ± 0.028) (n = 20, 1). SecOrL (including sinus), 0.08 – 0.10 (0.090 ± 0.007); SecOrW, 0.08 – 0.10 (0.089 ± 0.006) (n = 20, 1). OvL, 0.16 – 0.18 (0.167 ± 0.006); OvW, 0.18 – 0.22 (0.198 ± 0.011) (n = 13, 1). Longest AvRosL per zooid, 0.08 – 0.12 (0.096 ± 0.013) (n = 20, 1). Largest colony observed 7 × 4 mm.
Description
Colony forming an irregular, unilaminar, encrusting sheet; white in colour. Zooids ( Figure 15 View Figure 15 (a – c)) distinct, outlined by opposing columns of small areolae with a line of calcification between them; six or seven areolae along each lateral margin. Frontal wall convex; smooth or sparsely granulated. Primary orifice deeply immersed, broader than long, lyrula ( Figure 15 View Figure 15 (a)) ranging from tapering and truncate, to moderately broad and alate; condyles blunt, weakly denticulate at end, directed proximomedially. Peristome a raised collar, often tall, with a conspicuous, U-shaped secondary sinus ( Figure 15 View Figure 15 (a)); zooids often have a sharp, umbonate or conical, sometimes wing-like projection on peristomial rim on each side of groove. Zooids have three or four ephemeral distal oral spines, in about equal proportions. In zooids destined to produce an ovicell, distal margin of secondary orifice serrate ( Figure 15 View Figure 15 (a)). Ovicells typically widespread in colony, present in third or fourth zooid generation from ancestrula. Ovicell ( Figure 15 View Figure 15 (b, c)) hyperstomial; smooth, with conspicuous pseudopores except in proximocentral region, periphery covered with secondary calcification. Peristome extends across proximal edge of ooecium, giving rise to low or tall, conical or horn-like process on each side ( Figure 15 View Figure 15 (b)). Zooids lack a frontal avicularium (41%) or have a single one (59%) (n = 46) on either side proximal to orifice; rostrum strongly tilted to one side, upper rostral edge serrate; crossbar thin or lacking; mandible elongate, tapering, blunt, pointing proximally or angled slightly medially or laterally. Enlarged avicularia (not shown) occurred but were rare in our material; those observed were single, on either side of orifice, pointing proximally, nearly as long as zooid; rostrum sub-spatulate, widening distally, with coarsely toothed rim. Ancestrula ( Figure 15 View Figure 15 (d)) tatiform, with moderately wide proximal cryptocyst and nine marginal spines; first daughter zooid has five orificial spines.
Remarks
The original description and illustrations of Parasmittina serrula from Maui, Hawaiian archipelago ( Soule and Soule 1973), indicate small zooids (average ZL, 0.37 mm; ZW, 0.28 mm); a granulated frontal wall; usually four oral spines; a denticulate distal orificial margin; a narrow to medium-width, tapering, non-alate lyrula rather widely separated from the condyles; often tall projections on the peristome lateral to the sinus groove; small, usually paired frontal avicularia lateral or proximolateral to the orifice, with the rostrum pointing proximally, turned onto one side, and having the upper edge finely serrate; occasional giant avicularia replacing the smaller type, with coarsely serrate rostral margins; and a tatiform ancestrula with a wide proximal cryptocyst and eight marginal spines.
Most populations described subsequent to the original description have similarly small zooids, but vary in the range and mode of oral spine number, lyrula shape, the frequency of single versus paired lateral-oral avicularia, and the number of spines on the ancestrula. In fact, some of this variation may not be intraspecific, and some previous records of P. serrula may actually represent different species. For example, in nominal P. serrula from the Solomon Islands ( Tilbrook 2006), the lyrula is narrow, tapering, non-alate and relatively distant from the condyles (as expected in P. serrula ), but other characters differ from those in Hawaiian material: zooid size is much larger (ca. 0.65 × 0.40); the lateral-oral avicularia are longer, of somewhat different shape, scarcely elevated and not tilted as much laterally; the peristome is mildly asymmetrical and much thinner, without horn-like projections proximally and/or distally on the peristomial rim; and zooids show an additional, non-tilted type of small frontal avicularia. Material from Vanuatu ( Tilbrook et al. 2001) described as P. serrula likewise may not be that species. Judging from the SEM images, zooids are around the same size as in Hawaiian material, but the peristome is thin and mildly asymmetrical, and the lyrula is low, moderately broad and conspicuously alate. Finally, the identity of Philippine material (Scholz 1991) with P. serrula is questionable. Scholz did not provide measurements; the ancestrula has nine spines, as at Okinawa, and enlarged avicularia with a serrate margin occur, as is typical of P. serrula . However, zooids appear to lack tilted, single or paired lateral-oral avicularia but instead have non-tilted adventitious avicularia with a smooth rostral edge, situated in the midline proximal to the orifice; the shapes of the peristome and proximal sinus are unusual for P. serrula ; and there appear to be two or three oral spines. The lyrula is not evident in Scholz ’ s figures.
Our material differs from the original and most subsequent descriptions in that zooids either lack a lateral-oral avicularium or have only a single one, in about equal proportions. Additionally, enlarged avicularia were rare; we observed them in only a few zooids in colony NSMT-Te 1101. Finally, the pronounced, paired, horn-like projections on the distal peristomial rim have not been reported in other populations, though previous workers may have missed them in the frontal-view SEM images typically used for illustrations (e.g. see Dick et al. 2006, fig. 9(a)). As a further complication, among several specimens at Okinawa, colonies showed either of two distinct morphologies. In one ( Figure 15 View Figure 15 (a, b)), the zooids have a smooth frontal wall and the lyrula is moderately broad and weakly alate. In the other ( Figure 15 View Figure 15 (c)), the frontal wall is granulated (as is typical of P. serrula ) and the lyrula is narrower, tapering and either non-alate or weakly alate. Since other characters are similar in the two forms (small zooid size; three or four oral spines; lateral-oral avicularia single or lacking; paired, horn-like projections can occur both proximally and distally on the peristomial rim), we attribute the observed differences to intraspecific, inter-colony variation. However, the differences also mirror the situation in an Alaskan Rhynchozoon population ( Dick and Mawatari 2005) in which two morphological types with minor differences represented genetically divergent lineages and probably different species.
Occurrence
We found a total of eight colonies, at the SES and MIN sites ( Table 1). Nominal Parasmittina serrula is broadly distributed in the subtropical to tropical, central to western Pacific. It has previously been reported from Hawaii, the Solomon Islands, Vanuatu, the Philippines, Kermadek Ridge ( New Zealand) and the Great Barrier Reef (see the synonyms for references). There is also a curious disjunct record from Carrie Bow Cay ( Belize) in the Caribbean ( Winston 1984). The Caribbean material appears indistinguishable from Hawaiian material and may reflect anthropogenic dispersal on ships ’ hulls from the Pacific through the Panama Canal.
| SES- |
Southeastern Shanxi Teachers School |
| SES |
Southeastern Shanxi Teachers School |
| NHMUK |
Natural History Museum, London |
| MIN- |
University of Minnesota |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Celleporaria triangula Seo, 1994
| Dick, Matthew H. & Grischenko, Andrei V. 2016 |
Celleporaria triangula:
| Seo 2005: 397 |
Celleporaria triangula Seo, 1994 , p. 189
| , Seo 1994: 189 |