
Terebellides fauveli, Parapar & Martin & Moreira, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4771.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:A46FAF72-6F95-4DA3-A41D-FE770D6EDF1F |
|
DOI |
https://doi.org/10.5281/zenodo.3816141 |
|
persistent identifier |
https://treatment.plazi.org/id/03BF4239-FFE3-FFD2-EAF8-C0459C68F84D |
|
treatment provided by |
Plazi |
|
scientific name |
Terebellides fauveli |
| status |
sp. nov. |
Terebellides fauveli View in CoL sp. nov.
Figures 11B View FIGURE 11 , 12B View FIGURE 12 , 16–20 View FIGURE 16 View FIGURE 17 View FIGURE 18 View FIGURE 19 View FIGURE 20 , 37 View FIGURE 37 ; Tables 1, 2 urn:lsid:zoobank.org:act:F388A555-F6F0-48A8-93C5-C244DADDA76B
Material examined. Type material. Thirty-six specimens: holotype (NHMD-636920); thirty-five paratypes (NHMD-231435, NHMD-231442–231445, NHMD-231447, NHMD-231450–231452, NHMD-231461, NHMD- 231463, NHMD-636921 to NHMD-636922). Non-type material. Seven specimens ( MNCN 16.01 View Materials /18585 to 16.01/18590, Table 1) .
Diagnosis. Body of medium/large size (15–35 mm in length). Branchiae comma-shaped, with anterior lobe and posterior ventral lobes thinner than dorsal ones, directly emerging from branchial stem and with well-developed thin terminal filament. TC1 notopodia and notochaetae longer than following thoracic ones. Thoracic neurochaetae with rostrum / capitium length about 4/1, and capitium with a first row of 2–3 very large teeth followed by three rows of also large, but smaller teeth.
Description based on holotype
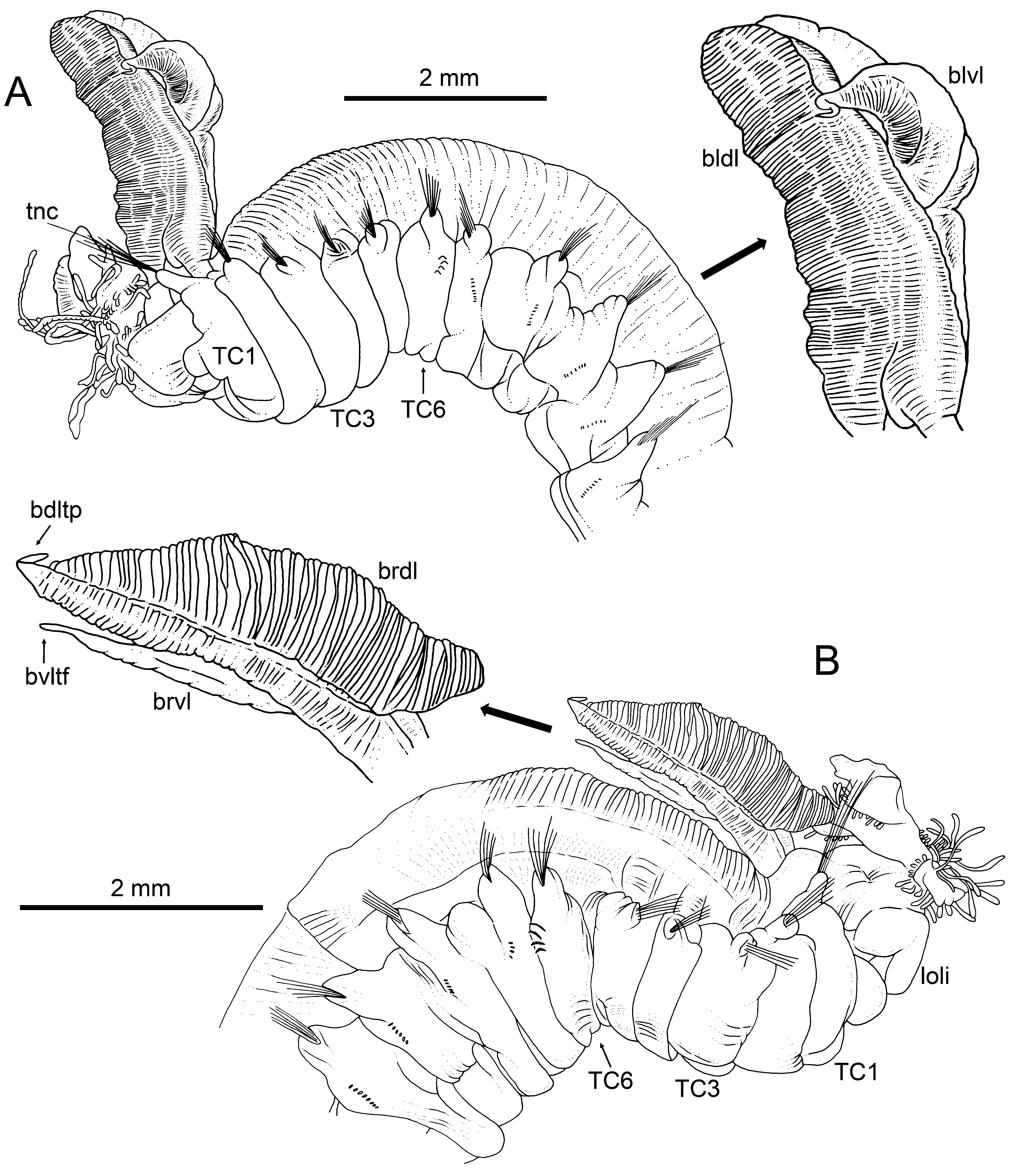
Measurements and general body features. Complete specimen, 30.0 mm long and 3.0 mm wide ( Fig. 12B View FIGURE 12 ). Body tapering posteriorly with segments increasingly shorter and crowded towards pygidium. Prostomium compact; large tentacular membrane surrounding mouth, with some typical buccal tentacles with expanded tips. SGI as expanded structure, below tentacular membrane ( Fig. 16 View FIGURE 16 B−C). Low lateral lappets on SGIII–VII (TC1–5) much larger in SGIII–V (TC1−3) and latero-dorsally white in TC3−4 ( Fig. 16 View FIGURE 16 A−B).
Branchiae. Branchiae comma-shaped, arising as single structure from SGIII, consisting of a single short stalked mid-dorsal branch with one pair of long dorsal (upper) unfused lobes, reaching TC6−7 ( Fig. 12B View FIGURE 12 , 16 View FIGURE 16 B−C), and one pair of ventral (lower) smaller and thinner lobes (about half length of dorsal), not fused together and emerging after a short fusion line of dorsal lobes ( Fig. 19 View FIGURE 19 A−B). Anterior projection of dorsal lobes (lobe 5) about 1/7 length of posterior dorsal lobes ( Fig. 16A, C View FIGURE 16 ). Pointed projection of posterior region only in lower lobes, as a thin but welldeveloped terminal filament ( Fig. 16D View FIGURE 16 , 17 View FIGURE 17 B−C, 19A−B). Both sides of branchial lamellae with rows of cilia ( Fig. 17 View FIGURE 17 B−C, 19C); ciliated tufts or papillae not seen.
Thorax. Eighteen pairs of notopodia (SGIII−XX), much more developed and with longer notochaetae in TC1 than in following ones ( Fig. 16 View FIGURE 16 C−D, 17A−B, 19D). All notochaetae as simple capillaries. Neuropodia as sessile pin- nules, from TC6 (SGVIII) to pygidium, with around 15 uncini in single rows starting from TC7 (SGIX) throughout. First neuropodia (TC6) with four sharply bent, acute tipped, geniculate chaetae ( Fig. 18A View FIGURE 18 ) having small minute teeth forming an ill-defined capitium ( Fig. 19 View FIGURE 19 E−F). From TC7, neuropodia with 8–14 uncini per torus in one row, with long shafted denticulate hooks, a very long rostrum about four times longer than capitium, with 2(3) very big teeth above main fang, surmounted by two rows of 3−4 also big, but smaller teeth and an upper crest of several smaller denticles ( Fig. 18 View FIGURE 18 B−C).
Abdomen. About 40% of body length, with 29 pairs of neuropodia as erect pinnules, with around 30 uncini per torus having three teeth above main fang surmounted by an upper crest of several minute teeth ( Fig. 18D View FIGURE 18 ).
Other body features. One nephridial papilla behind each TC1 notopodium and nephridial openings in TC4 and TC5 notopodia. Pygidium blunt, funnel-like depression.
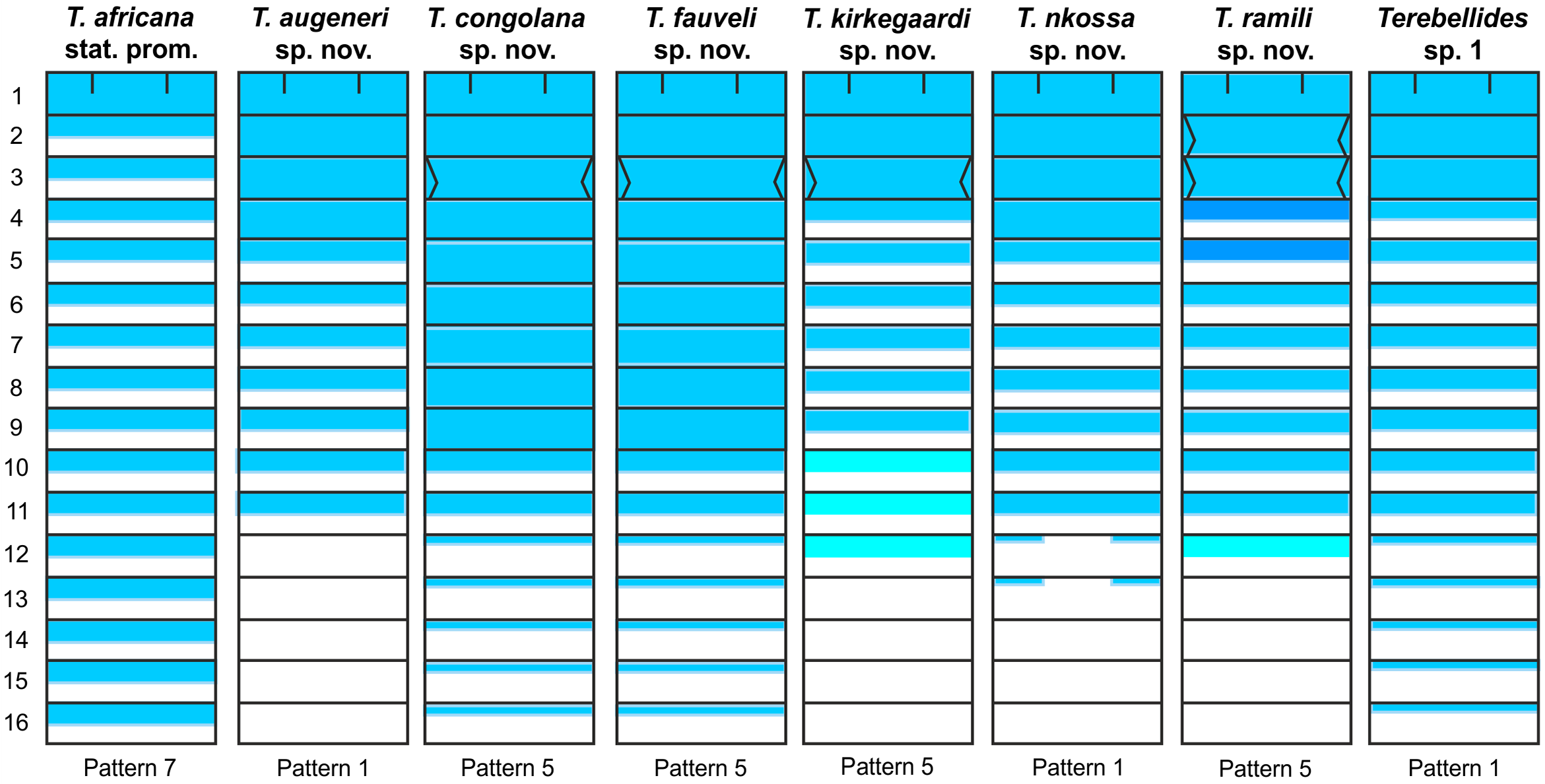
Methyl green staining pattern. Anterior CH 1 to CH 9 solid; CH 10 to CH 18 striped; being CH 10 to CH 11 much more marked than following; J-shaped glandular region marked lateral to CH 3; near pattern 5 of Schüller & Hutchings (2010) ( Fig. 37 View FIGURE 37 ).
Variability. Body between 15 and 35 mm in length.
Type locality. Ivory Coast; 80–90 m depth (Table 1) .
Distribution and bathymetry. Widely distributed along the West African coasts, from Liberia to South Angola; 42−200 m depth ( Fig. 11B View FIGURE 11 ; Table 1).
Etymology. The species is named after the French zoologist Pierre Fauvel (1866–1958) for his many contributions to the study of the Polychaeta across world oceans, especially in Europe and Africa.
Remarks. Three Terebellides species have been previously described with TC1 provided with large notopodia and long notochaetae: T. kobei Hessle, 1917 ( Japan) , T. californica Williams, 1984 (California) , T. mediterranea , and two in this work: T. congolana sp. nov. and T. fauveli sp. nov.
Terebellides fauveli sp. nov. has shorter TC1 notopodia and notochaetae than T. kobei , lacks a conspicuous triangular dorsal projection in TC3 notopodia (present in T. kobei ), has a white lateral colouration in anterior chaetigers (absent in T. kobei ) and has a different capitium denticulation, being all teeth small and of similar size in T. kobei , as illustrated by Hessle (1917: textfig. 32, p. 140).
Terebellides californica was described from shelf and slope depths in the Pacific Ocean (Oregon to Western Mexico) and is characterized by the author only by the “ very well developed first chaetiger with greatly prolonged fine notosetae ” ( Williams, 1984: p. 128), which is clearly longer than the one in T. fauveli sp. nov. and lacks any special development in TC3 and body colour pattern. This species was later redescribed by Hilbig (2000) as having a “ trilobed structure of the peristomium ” as potential new diagnostic character, which is not present in T. fauveli sp. nov. However, in our opinion, the relevance of this character should be considered with caution as its presence in T. californica has not been sufficiently assessed.
Terebellides mediterranea most closely resembles T. fauveli sp. nov. in branchial shape. However, T. fauveli sp. nov. bears larger branchiae that reach TC6 (instead of TC3−4) and has posterior branchial lobes with terminal filament. Moreover, both species differ in number and size of the denticulation of thoracic uncini, with 3−4 mediumsized teeth above main fang and surmounted by a row of 6−7 slightly shorter tooth in T. mediterranea (see Parapar et al., 2013: Fig. 7B View FIGURE 7 ).
Terebellides anguicomus and T. lanai Solís-Weiss et al., 1991 resemble T. fauveli sp. nov. in the shape (commalike) and size of the branchiae, also reaching TC6. However, T. lanai bears branchial lobes fused for most of their length, and T. anguicomus has a large transverse dorsal hump on TC5, 17 thoracic chaetigers (instead of 18) and a shorter TC1.
While some of the French species recently described by Lavesque et al. (2019), such as T. lilasae Lavesque, Hutchings, Daffe, Nygren & Londoño-Mesa, 2019 , have similar general branchial shape as in T. fauveli sp. nov., the capitium denticulation of the thoracic uncini is very different; in fact, the shape of the thoracic uncini in T. fauveli sp. nov. has not been recorded so far in any other species of the genus.
Among the West African species, T. fauveli sp. nov. apparently presents the widest bathymetric and geographic distribution. Non-type specimens collected off Congo (N’Kossa gas field, Table 1) show subtle differences in the shape and size of the terminal filament of lower branchial lobes, size of TC1 and thoracic uncini teeth (cfr. Fig. 17−18 View FIGURE 17 View FIGURE 18 vs. 19−20). Therefore, we cannot discard they may represent a different, close taxon. More extensive sam- pling across West Africa and further morphological and molecular analyses may help to test the present wide distribution of T. fauveli sp. nov. and the possible existence of several cryptic species.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.