
Sphenomorphus melanopogon ( Duméril & Bibron 1839 ), Dumeril & Bibron, 1839
|
publication ID |
https://doi.org/10.11646/zootaxa.3490.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:A0DB5E47-3CBC-44D7-83A1-5235E9EE289D |
|
DOI |
https://doi.org/10.5281/zenodo.5874678 |
|
persistent identifier |
https://treatment.plazi.org/id/03BF87DA-FFBA-FF90-99E9-75A3FD65E2A6 |
|
treatment provided by |
Plazi |
|
scientific name |
Sphenomorphus melanopogon ( Duméril & Bibron 1839 ) |
| status |
|
Sphenomorphus melanopogon ( Duméril & Bibron 1839)
Lygosoma melanopogon Duméril and Bibron 1839: 723 .
Lectotype: MNHN 1245 (designated herein, and according with previous restrictions on the type series by Guibé 1954); type locality Nouvelle Hollande (in error for Timor).
Scincus naevius Duméril and Bibron 1839: 724 . (Junior objective synonym made available by subsequent recognition as a valid taxon prior to 1961). Lectotype: MNHN 1245, designated herein; type locality Nouvelle-Hollande (in error for Timor).
Lygosoma florense Weber 1891: 173 , plate 14, figs 2–3. Lectotype: ZMA 11060, designated herein; restricted type locality (due to lectotype selection) Maumere, Flores.
Lygosoma ( Hinulia) Kühnei Roux 1910: 237 , plate 13, fig. 2. Lectotype: NHMB 5514 (designated by Kramer 1979); type locality Kei Islands [= Kepulauan Kai].
Sphenomorphus florensis nitidus Dunn 1927: 5 . Holotype: AMNH 32068; type locality Komodo.
Sphenomorphus florensis barbouri Dunn 1927: 5 . Holotype: AMNH 32203; type locality north coast of Wetar, near Uhak.
Sphenomorphus florensis weberi Dunn 1927: 6 . Holotype: MCZ 20996 [erroneously cited as 2099 in description]; type locality Damma Island [= Pulau Damar].
Sphenomorphus kuehnei Kramer 1979: 163 .
Emendation of Kühnei Roux (Article 32.5.2.1 of the Code of Zoological Nomenclature).
Diagnosis. This species can be readily differentiated from the majority of Sphenomorphus species by the combination of finely striate dorsal scales, the imbricate scales on the dorsal surface of the pes extending onto the plantar surface between the fourth and fifth digits, and three or more supraoculars contacting the frontal. It can be further differentiated from the few Sphenomorphus (all members of the variegatus group) with all of these character states by the combination of smoothly rounded subdigital lamellae, presence of auricular lobules and the postmental modally only contacting a single infralabial on each side. When present, the black throat is also a useful diagnostic character.
Distribution. Known from the Lesser Sunda Islands, specifically from west to east: Pulau Deli, Pulau Tinjil, Sumbawa, Sumba, Komodo, Padar, Flores, Pulau Adonara, Pulau Semau, Timor, Pulau Alor, Wetar, Pulau Babar, Pulau Damar, Pulau Teun, Pulau Banda Besar and Kepulauan Kei ( Fig. 9 View FIGURE 9 ). Auffenberg (1980) also lists the species from Pulau Rinca and Pulau Besar, although I am unaware of any specimen-based records to support this. Although the type series of Lygosoma Kühnei was purportedly collected from the Kei Islands, there are no other records of the species from this easternmost extremity of the species' distribution. The monograph by Roux (1910) is largely based on material collected by Jean Roux and his colleague Hugo Merton in the Kei and Aru Islands in 1907–08. However, two species, Lygosoma Kühnei and the agamid Lophognathus temporalis Günther , were not collected by Roux and Merton, but were reported solely on the basis of specimens purchased from the natural history dealer Hermann Rolle in Berlin in 1905, with the specimens said to have been collected by the late Herrn Kühne. Rolle's Kühne is presumably the ornithological and entomological collector Heinrich Kühn ( 8.ii.1860 Erlbach– 26.vii.1906 Surabaya), who lived at Toeal [= Tual] in the Kei Islands for several years, but who also collected extensively in the surrounding islands, including Damar (1898), Banda Besar (1898), Wetar (1902) and Baber (1905) ( Hartert, 1900a – b, 1901, 1904, 1906, 1907), all of which are within the confirmed distribution of S. melanopogon . There are specimens of Sphenomorphus melanopogon purchased from H. Rolle in 1900 in the BMNH collection (BMNH 1900.2.9.5–10) which bear the locality Damar I., a locality from which S. melanopogon was independently collected by the Siboga expedition ( Dunn, 1927). These are likely to have been collected by Kühn. Lophognathus temporalis has also been reported from Damar and Baber ( de Rooij, 1915), localities visited by Kühn. Hence it is possible that the type locality for L. Kühnei is incorrect.
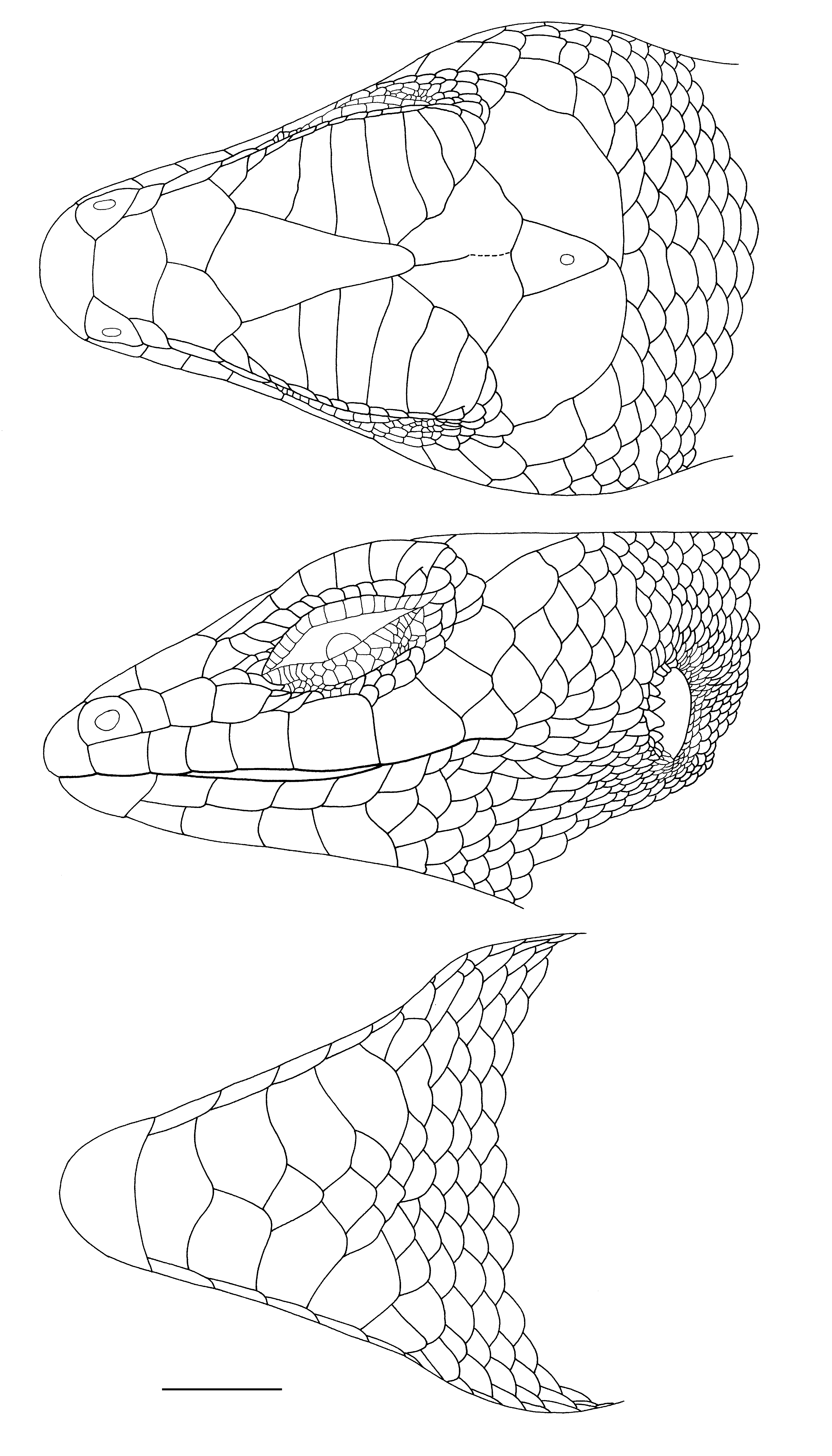
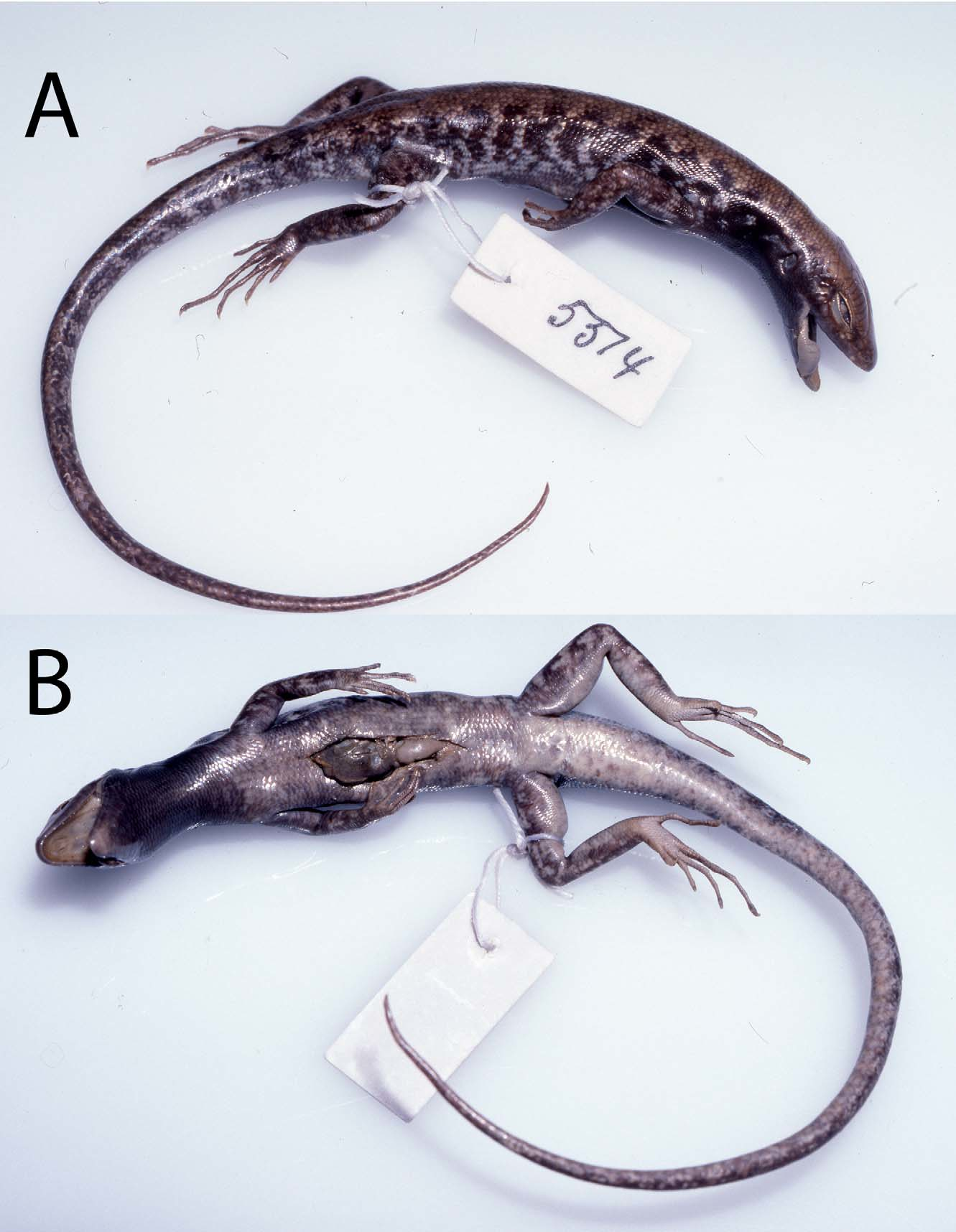
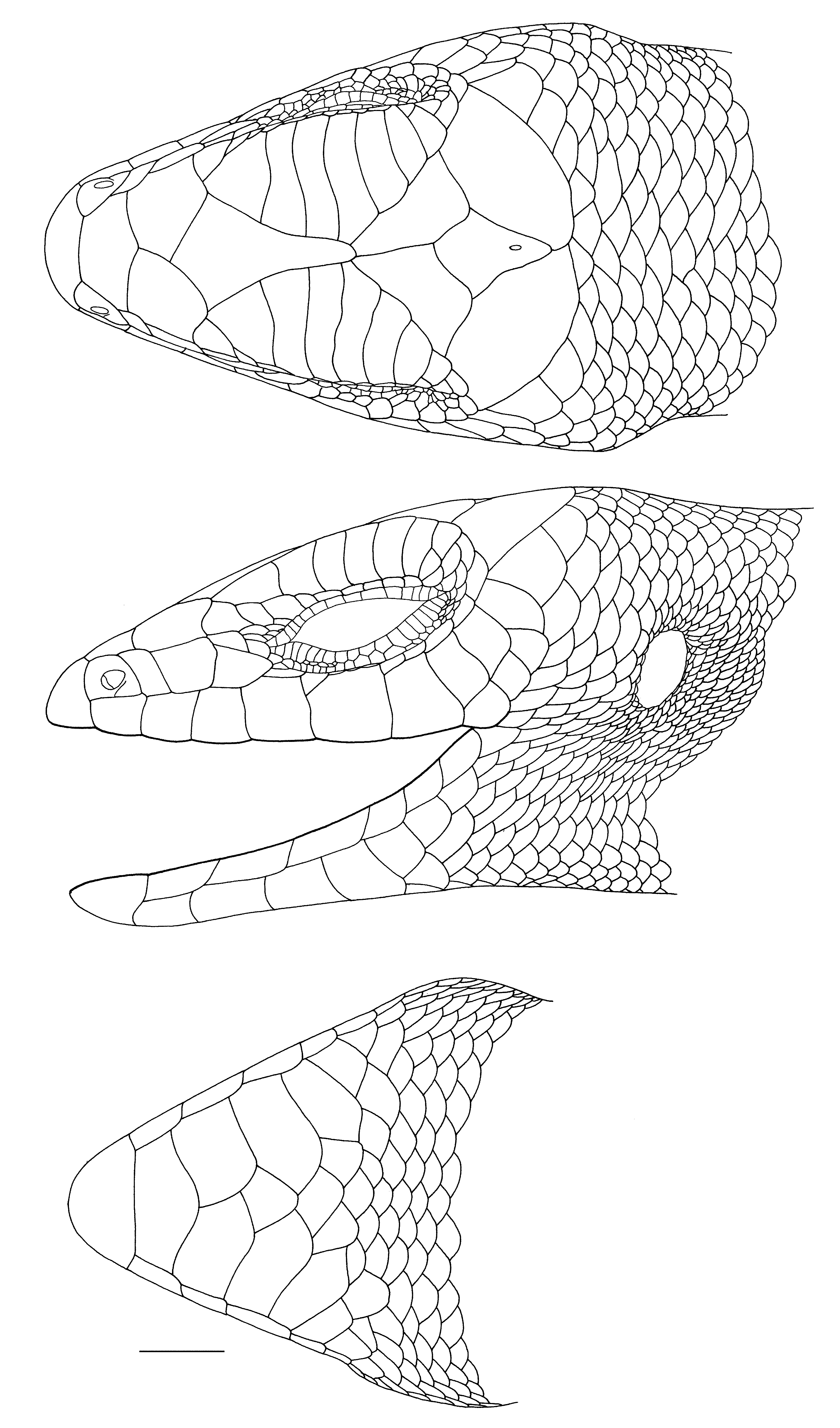
Redescriptions of Lectotypes. The lectotype of Lygosoma melanopogon (MNHN 1245; Figs. 1A View FIGURE 1. A –B, 2) has the following character states: dorsal lobe of rostral with a slightly concave posterior border; nasals broadly separated; nostril central in nasal; supranasals absent; prefrontals in broad contact; supraoculars seven, first four in contact with frontal, last four in contact with frontoparietal; supraciliaries 14/14, first separated from frontal; frontoparietals in broad median contact; parietal eye spot at junction of second third and last third of interparietal length; parietals in broad contact behind interparietal; anterior loreal taller than wide and taller than posterior loreal; posterior loreal squarish, a little wider than tall, and narrowest ventrally; presuboculars two, first largest, last interdigitating with supralabials; suboculars 4/3; postsuboculars 7/8, first interdigitating with supralabials; supralabials seven, fifth below centre of eye, last two each divided into an anteroventral and a posterodorsal scale; primary temporals two, anterodorsal and posteroventral, the more dorsal very small; lower secondary temporal single, overlapping upper secondary temporal; scales bordering parietals between upper secondary temporals eight; ear with six low rounded lobules along anterior margin; infralabials 8/7, only first contacting postmental; first pair of chin shields in median contact; second pair medially separated by a single scale; third pair of chin shields separated by three scales; all chin shields laterally contacting infralabials.
Longitudinal scale rows at midbody 43; scales in a paravertebral row (A) 71, not broader than adjacent dorsal scales; dorsal scales finely striate, laterally and ventrally becoming smooth; median pair of precloacal scales enlarged, laterally overlapping adjacent precloacals; lamellae below fourth toe (A) 30/29; scales above fourth toe four at level of metatarsophalangeal joint, decreasing distally so that last two scales single.
SVL 60 mm; AGL 27 mm; tail regenerated; FLL 22 mm; HLL 37 mm; HL 13.1 mm; HW 9.5 mm; HL 7.6 mm.
The male lectotype of Lygosoma florense (ZMA 11060; Figs. 4–5 View FIGURE 4. A View FIGURE 5 ) has the following differences in scalation and morphometrics from those described for the lectotype of L. melanopogon : posteriormost supraocular does not contact frontoparietals; supraciliaries 13/13; frontoparietals partly fused posteriorly; parietal eye spot slightly posterior to centre of interparietal; posteriormost presubocular barely interdigitating with supralabials; suboculars 5/4; postsuboculars 8/6; lower secondary temporal overlapped by upper secondary temporal (overlap pattern as for the lectotype of L. melanopogon in remainder of type series); ear with seven conical lobules along anterior margin; infralabials 8/9.
Longitudinal scale rows at midbody 46; paravertebral scales (A) 84; subdigital lamellae (A) 30/34.
SVL 68 mm; AGL 32 mm; tail regenerated; FLL 21.5 mm; HLL 37.5 mm; HL 15.8 mm; HW 11.4 mm; HD 9.0 mm.
The male lectotype of Lygosoma Kühnei ( Figs. 6–7 View FIGURE 6. A View FIGURE 7 ) has the following scalational and morphometric characteristics that differ from those described for the lectotype of L. melanopogon : supraciliaries 12/13; anterior loreal about same height as posterior loreal; suboculars 3/3; postsuboculars 5/6; lower secondary temporals two, the upper of the two including the upper portion of the primary temporal; ear with five low rounded lobules along anterior margin; infralabials 7 (on left — right side not counted).
Longitudinal scale rows at midbody 40; paravertebral scales (B) 77; subdigital lamellae (B) 31.
SVL 66 mm; TL 120 mm; FLL 24 mm; HLL 39 mm. AGL and head measurements not taken.
A new scalational character of the foot in some Sphenomorphus species
Within Sphenomorphus , there is a group that is recognisable on the basis of a character in the pes that is unique in skinks. The character concerns the squamation on the postaxial edge of the fourth metatarsal, that is, the posterior edge of the sole of the foot between the level of the free part of the fourth digit and the base of the fifth digit.
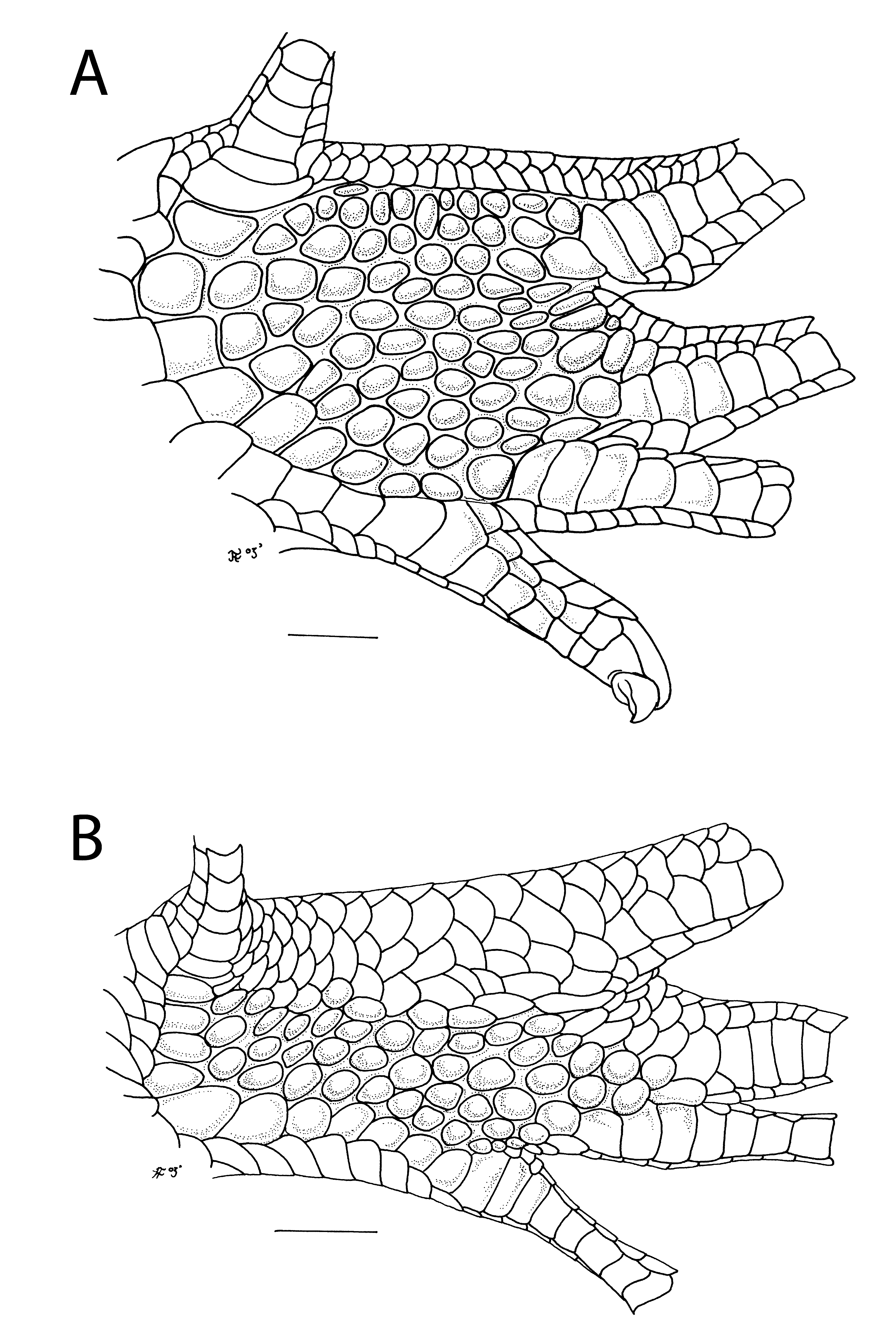
In the primitive condition, as seen in all other skinks, including the generally plesiomorphic scincine genus Plestiodon , there is a distinct line of demarcation between the field of flat, imbricate scales on the dorsal side of the pes and the field of more rounded or tuberculate, non-imbricate scales of the ventral side. In the derived condition, this line of demarcation has shifted onto the ventral surface and has become less distinct; in other words, it looks as if the imbricate scales of the dorsal surface have extended onto the ventral surface with a gradual merging of the morphology of the scales in the two fields ( Fig. 15 View FIGURE 15 ). The extent of this ventral extension can vary from just on the edge of the ventral surface as in S. multisquamatus to almost the entire ventral surface, leaving just a single arcuate line of juxtaposed scales between the bases of the first and fifth digits, as in S. anomalopus .
The derived character state has been obliquely mentioned by Smith (1935) and Taylor (1963) for S. maculatus , although both authors emphasised the restriction of the tuberculate juxtaposed scales rather than the extension of the imbricate scales. It has also been illustrated for S. anomalopus by Smith (1937).
Species which show the derived character state are S. amabilis (Müller) ( MCZ 112192), S. anomalopus (Boulenger) ( BMNH 1913.6.4.3), S. cyanolaemus Inger and Hosmer ( MCZ 83210–11), S. dussumieri (Duméril & Bibron) ( AMNH 107258, BMNH 1946.8.15.38–44), S. maculatus (Blyth) (AM R171873–74), S. melanopogon , S. multisquamatus Inger ( AMNH 92893–94, 111913, MCZ 83213), S. nigrilabris (Günther) ( BMNH 1946.8.19.69, MCZ 110309), S. sabanus Inger ( MCZ 22951), S. sanctus (Duméril & Bibron) ( MCZ 7663a–b), S. sarasinorum (Boulenger) ( BMNH 1946.8.15.9–10), S. simus (Sauvage) (AM R 120338, R 120351), S. striolatus ( ZMA 10945), S. tersus (Smith) ( BMNH 1946.8.3.22–23, MCZ 39285), S. tropidonotus (Boulenger) ( SMF 14472; AMNH 142999) and S. variegatus ( MCZ 20138, 26610).
A sharp line of demarcation between imbricate dorsal scales and juxtaposed plantar tubercles (the plesiomorphic state) occurs in S. acutus (Peters) (AM R 120570 –71, R104881, R104886, MCZ 20115; USNM 497028), S. annectens (Boulenger) ( MSNG 29115), S. buettikoferi (Lidth de Jeude) ( RMNH 4471a–c), S. cameronicus Smith ( BMNH 1946.8.3.27), S. concinnatus (Boulenger) (AM R 121595, R 121764), S. consobrinus (Peters & Doria) ( SMF 14455, 14457, 14459–60), S. courcyanus (Annandale) ( BMNH 1946.8.16.91), S. cryptotis Darevsky, Orlov and Ho ( VNMN 1225–26, 1288), S. darlingtoni (Loveridge) (AM R23019–20), S. diwata Brown and Rabor ( USNM 496805), S. formosensis Van Denburgh ( AMNH 34878–79, 34887), S. fragosus Greer and Parker ( MCZ 92262–65), S. granulatus (Boulenger) (AM R129754, R129772), S. incognitus (Thompson) ( AMNH 33178–79, 34904, MCZ 110307, USNM 65368), S. indicus (Gray) (AM R96605, R171635–36, R173604, MCZ 39263–64), S. jobiensis (AM R24721, R24789), S. kinabaluensis (Bartlett) ( MCZ 22952, 43496), S. maculicollus Bacon ( FMNH 161484), S. meyeri (AM R118736, R118739), S. mimicus Taylor ( MCZ 39256), S. mimikanus (Boulenger) ( BMNH 1946.8.15.11), S. muelleri (Schlegel) (AM R13865, R14569), S. murudensis Smith ( BMNH 1946.8.15.6), S. neuhaussi (Vogt) (AM R9867–68), S. praesignis (Boulenger) ( BMNH 1906.2.28.15, 1916.3.27.22, 1946.8.15.53, 1974.3857, 1974.4988), S. pratti (Boulenger) (AM R129522, R129682), S. scotophilus (Boulenger) (AM R153110–11, BMNH 1946.8.16.94), S. stellatus (Boulenger) ( MCZ 39283), S. taiwanensis Chen and Lue (AM R 161050, R 161053), S. tanneri Greer and Parker (AM R31132–33, R 121594), S. taylori Burt ( MCZ 65857, 96223), S. temmincki (Duméril & Bibron) (AM R6809–10; USNM 43346), S. textus (Müller) ( BMNH 95.2.27.2), S. transversus Greer and Parker ( MCZ 76485), S. wollastoni (Boulenger) ( BMNH 1946.8.3.70), S. woodfordii (Boulenger) (AM R91266, R93032), and all members of the S. maindroni (Sauvage) species-group of Greer and Shea (2004). This condition is also present in a number of Philippine species formerly in Sphenomorphus that were recently transferred to other genera by Linkem et al. (2011), including Insulasaurus arborens (Taylor) ( MCZ 20142–43, USNM 496765), I. victoria Taylor ( MCZ 44162), I. wrighti (Taylor) ( MCZ 26301), Otosaurus cumingi Gray ( MCZ 26302; USNM 229156), Parvoscincus beyeri (Taylor) ( MCZ 26365; USNM 337768), Pa. decipiens (Boulenger) ( MCZ 20144–45), Pa. laterimaculatus (Brown & Alcala) ( USNM 318341, 318343–44), Pa. lawtoni (Brown & Alcala) ( MCZ 26375; USNM 318981), Pa. luzonensis (Boulenger) ( AMNH 67098, MCZ 163156, USMM 512761), Pa. steerei (Stejneger) ( MCZ 26409). Pinoyscincus coxi (Taylor) ( MCZ 26330; USNM 318437), Pi. jagori jagori (Peters) ( MCZ 20136–37, 26303, 173396), Pi. jagori divergens (Taylor) ( MCZ 20134), Tytthoscincus atrigularis (Stejneger) ( MCZ 26379), T. biparietalis Taylor ( MCZ 26385), and T. hallieri (Lidth de Jeude) ( MCZ 22632),
I can only speculate as to the functional significance of this distinctive morphology. In general, the skinks with the derived morphology tend to be gracile with relatively long legs. This suggests that they may be able to run more rapidly or to climb more adeptly than their shorter-legged relatives. Furthermore, the partial loss of the distinct plantar scale morphology suggests that during locomotion, the posterior edge of the plantar area of the pes may make less areal or temporal contact with the ground than in other skinks. Perhaps during high-speed locomotion or in climbing, these skinks deploy the foot in a way that lessens the contact of this part of the pes with the substrate.
In a recent genetic analysis of Philippine Sphenomorphus ( Linkem et al. 2011) , all five of the species showing the derived condition that were included in the analysis ( S. cyanolaemus , S. maculatus , S. multisquamatus , S. sabanus and S. variegatus ), together with one species not showing this condition ( S. indicus ) formed a distinct well-supported lineage ("Clade B"). Assuming that the derived condition represents a synapomorphy of part or all of this clade, with S. melanopogon also a member, the name Sphenomorphus can be applied to this clade. However, determination of the extent of Sphenomorphus in this restricted sense requires assessment of the character in the many species of Sphenomorphus that have not yet been examined for it, as well as genetic data for S. melanopogon and wider genetic sampling of non-Philippine taxa.
Specimens examined
Sphenomorphus melanopogon (n = 195) (all localities are in Indonesia)
Pulau Deli ( 7o00'S 105o32'E): SMF 53924–35 GoogleMaps . Sumbawa: SMF 23263–64, Sumbawa Besar ( 8o30'S 117o26'E) GoogleMaps ; 23265, Batoe-Doelang (= Batudulang 8o36'S 117o19'E) GoogleMaps ; 23266, Dompoe (= Dompu 8o32'S 118o28'E) GoogleMaps . Komodo: AMNH 32068 ( holotype), 32055–67, 32070 ( paratypes), 2000'; 32101–04 ( paratypes of nitidus), sea level ; SMF 61005–06 . Pulau Padar ( 8o40'S 119o35'E): AMNH 32000–01, 32003, MCZ 27015, SMF 61015–16 GoogleMaps . Flores: BMNH 1946.8.15.50 ( paralectotype of florensis ), Maumeri [= Maumere 8o37'S 122o14'E] GoogleMaps ; BMNH 97.6.21.18–20 ; BMNH 97.12.30.74, E. Flores ; MCZ 9319, Larutuka [= Larantuka 8o21'S 122o59'E] GoogleMaps ; RMNH. RENA 5817a–b, Larutoko, E. Flores [=Larantuka] ; SMF 23267–69, Rana Mese [ 20 km SE Ruteng fide Mertens, 1930:140] ; SMF 23270–71, Endeh [= Ende 8o50'S 121o39'E] GoogleMaps ; SMF 23272–73, Wolo Waroe [= Wolowaru 8o46'S 121o54'E] GoogleMaps ; SMF 23283, Sita [ 8o29'S 120o14'E] GoogleMaps ; ZMA 11056–58 ( paralectotypes of florensis ), 11813a–c, Sikka, E. Flores [= Sika 8o45'S 122o12'E] GoogleMaps ; ZMA 11060 ( lectotype), 11059, 11061–64 ( paralectotypes of florensis ), Maumeri [= Maumere] ; ZMA 11814, Rieng Kemie [= Riangkamie 8o16'S 122o58'E] GoogleMaps ; ZMA 11815, Rioeng region, North coast [= Riung 8o28'S 121o03'E] GoogleMaps ; ZMA 11816a–p, Larantoeka [= Larantuka] ; ZMA 11817, Flimandiri, above Tabali, E. Flores [= Ili Mandiri 8o19'S 122o58'E] GoogleMaps ; ZMA 11818a–b, north slope of Flimandiri, E. Flores [= Ili Mandiri] ; ZMA 11819, no specific locality . Pulau Adonara ( 8o20'S 123o10'E): RMNH. RENA 4880a–b GoogleMaps . Pulau Semau ( 10o13'S 123o22'E): RMNH. RENA 2534a–l, Samao GoogleMaps . Timor: BMNH 1926.10.30.58–59, MCZ 25381–85, Lelogama, S. Timor [ 9o44'S 123o57'E] GoogleMaps ; MNHN 1245, 7110 ( lectotype, paralectotype of melanopogon ) ; RMNH. RENA 5817a–b, Noil Toko, Dutch Timor [=Noeltoko 9o33'S 124o22'E] GoogleMaps ; RMNH. RENA 2535a–k . Pulau Alor ( 8o15'S 124o45'E): BMNH 97.12.30.11–14 GoogleMaps . Wetar: AMNH 32203 ( holotype), 32192–202 ( paratypes), MCZ 27026–31 ( paratypes of barbouri) , nr Uhak , N coast [ 7o35'S 126o30'E]; NMBA 6476–77 GoogleMaps , RMNH. RENA 5532a–d ; SMF 14415–21, Iliwaki [= Ilwaki 7o56'S 126o26'E] GoogleMaps ; ZSM 20/1915a–b, Fliwaki [= Ilwaki] . Pulau Damar ( 7o09'S 128o40'E): MCZ 20996 ( holotype of weberi), Damma GoogleMaps ; BMNH 92.4.13.1–2, 1900.2.9.5–10, Damna Island . Pulau Teun ( 6o59'S 129o08'E): RMNH. RENA 5090a–c GoogleMaps ; BMNH 1932.1.1.1, Teoen, Damnar I., Banda See . Kepulauan Babar: RMNH. RENA 5089a–d, Teboe, Baber Eil. [= Tepa, Pulau Babar 7o52'S 129o31'E] GoogleMaps ; 7232a–b, Babber Is. ; ZMA unregistered (n = 6), Babar Eil. ; ZMA unregistered (n = 2), Tepa, Babar Eil . Pulau Banda Besar: RMNH. RENA 5091a–b , Lonkoer, Banda Is. [= Lonthoir ( 4o32'S 129o52'E). Kepulauan Kai : MCZ 27945 (paralectotype), GoogleMaps NMBA 5514 (lectotype) GoogleMaps , 5513, 5515–17 GoogleMaps ( paralectotypes of kühnei), Kei-Inseln.



| MCZ |
Museum of Comparative Zoology |
| AMNH |
American Museum of Natural History |
| ZMA |
Universiteit van Amsterdam, Zoologisch Museum |
| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| MSNG |
Museo Civico di Storia Naturale di Genova 'Giacomo Doria' |
| RMNH |
National Museum of Natural History, Naturalis |
| VNMN |
Vietnam National Museum of Nature |
| FMNH |
Field Museum of Natural History |
| MNHN |
Museum National d'Histoire Naturelle |
| ZSM |
Bavarian State Collection of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Sphenomorphus melanopogon ( Duméril & Bibron 1839 )
| Shea, Glenn M. 2012 |
Sphenomorphus kuehnei
| Kramer 1979: 163 |
Sphenomorphus florensis nitidus
| Dunn 1927: 5 |
Sphenomorphus florensis barbouri
| Dunn 1927: 5 |
Sphenomorphus florensis weberi
| Dunn 1927: 6 |
Lygosoma ( Hinulia ) Kühnei
| Roux 1910: 237 |
Lygosoma florense
| Weber 1891: 173 |
Lygosoma melanopogon Duméril and Bibron 1839: 723
| Dumeril 1839: 723 |
Scincus naevius Duméril and Bibron 1839: 724
| Dumeril 1839: 724 |