
Mystacodon selenensis, Lambert, Martinez-Caceres, Bianucci, Di Celma, Salas-Gismondi, Steurbaut, Urbina & Muizon, 2017
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2019v41a11 |
|
publication LSID |
urn:lsid:zoobank.org:pub:10C821A5-943E-41A0-8FFF-5D5169223126 |
|
DOI |
https://doi.org/10.5281/zenodo.3704892 |
|
persistent identifier |
https://treatment.plazi.org/id/03BFC706-2045-0965-FEE4-FBF08FACF84E |
|
treatment provided by |
Valdenar |
|
scientific name |
Mystacodon selenensis |
| status |
|
Lambert, Martínez-Cáceres, Bianucci, Di Celma, Salas- Gismondi, Steurbaut, Urbina & Muizon, 2017
HOLOTYPE. — MUSM 1917, a partial skeleton. Associated elements include the skull, missing most of the basicranium and the right squamosal and jugal; a fragmentary left tympanic; both dentaries; thyrohyal; cervical and thoracic vertebrae; ribs; sternum (four elements, including manubrium and xiphisternum); partial right and left forelimbs; and left innominate.
ETYMOLOGY. — From Selene, the Greek goddess of the moon, in reference to the Playa Media Luna, the type locality.
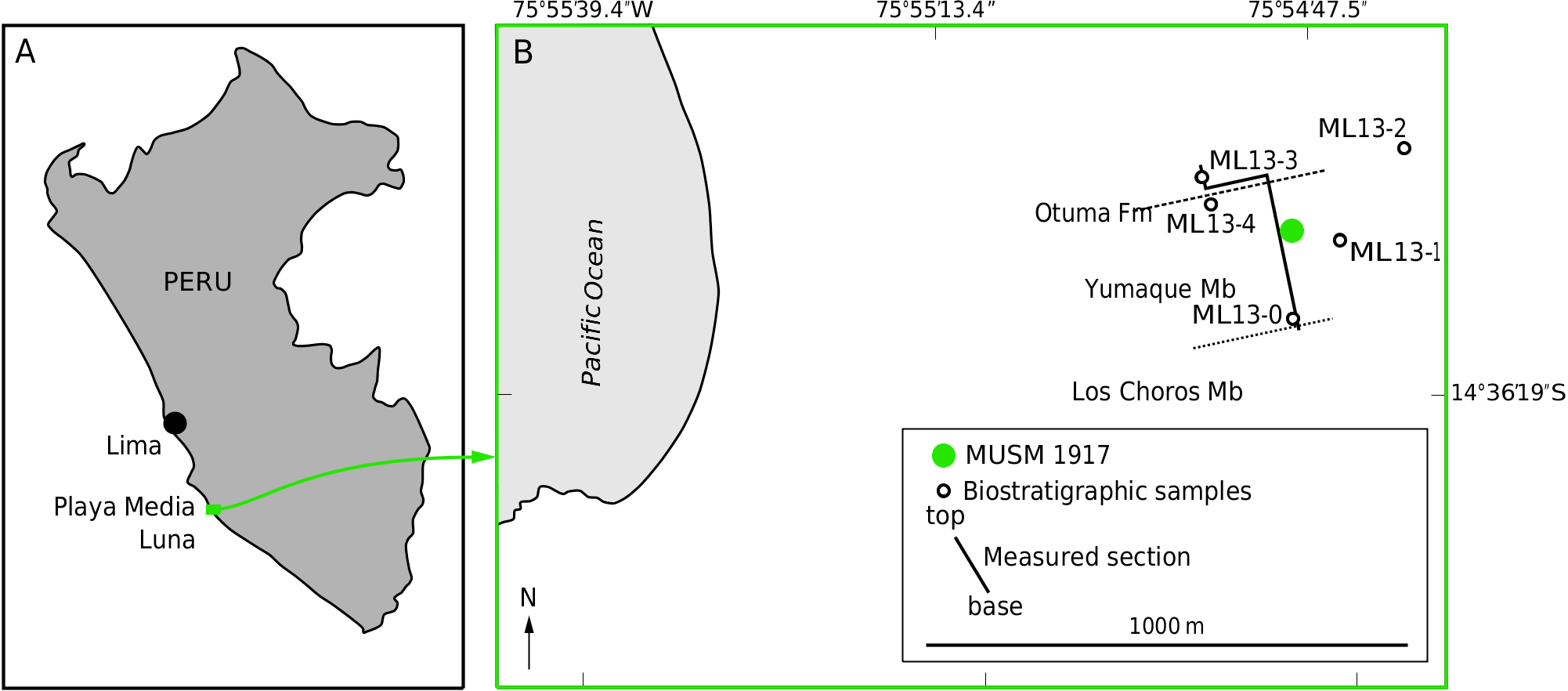
TYPE LOCALITY. — The type specimen comes from the Playa Media Luna (14°36’7.2”S, 75°54’48W), close to the Pacific coast in the southern part of the Pisco Basin, at about the level of the km 400 of the South Panamerican Highway ( Fig. 1 View FIG ).
TYPE HORIZON AND AGE. — The holotype of Mystacodon selenensis MUSM 1917 was discovered in the middle part of the Yumaque Member of the Paracas Formation, 77 m above the contact with the Los Choros Member ( Figs 2 View FIG ; 3 View FIG ) (see DeVries 2017 for a revised interpretation of the Eocene depositional sequence in the area). Layers of the Yumaque Member comprise finely laminated to massive, diatomacetous siltstones, containing pelagic microfossils, thin-shelled pectinid bivalves, and numerous fish scales; they represent deposition in distal (outer shelf), low-energy marine settings ( Lambert et al. 2017a; DeVries 2017). The calcareous nannofossil investigation of 37 sediment samples from the Yumaque Member and the overlying lower part of the Otuma Formation, taken along the stratigraphic section in the type locality of Playa Media Luna, allowed positioning the type horizon in the lower part of calcareous nannofossil zone NP19/20 of Martini (1971); based on age estimations by Agnini et al. (2014), an age of 36.4 Ma (early Priabonian, early late Eocene) has been proposed (see Lambert et al. 2017a for more details).
EMENDED DIAGNOSIS. — Mystacodon selenensis is identified as a Neoceti based on the following derived characters, absent in basilosaurid archaeocetes: partly open mesorostral groove; anteroposteriorly elongated rostral portion of maxilla; presence of an antorbital process of the maxilla; supraoccipital shield anterodorsally inclined; and distal epiphysis of the humerus divided in two angled radial and ulnar facets.
Mystacodon selenensis is referred to the Mysticeti due to the following combination of derived characters: dorsoventrally thin posterolateral region of maxilla on rostrum; antorbital process of maxilla separated from lacrimal; presence of a maxillary infraorbital plate; apex of zygomatic process of squamosal closely apposed to postorbital process of frontal or situated ventral to the latter; external occipital crest restricted to anterodorsal half of supraoccipital shield; and triangular supraoccipital shield.
Mystacodon selenensis differs from all other toothed mysticetes in bearing the following combination of features: 1) elongated nasal, the nasal being longer than the length of the frontal + parietal, at midline; 2) anteriorly located antorbital foramina, at about the level of the diastema between P2 and P3; 3) absence of real antorbital notch; 4) long jugo-squamosal suture, with an anterior prolongation of the zygomatic process of the squamosal, extending as far as the level of the postorbital process of the frontal anteriorly; 5) short anterior process of the frontal separating the posterior apices of the nasals; 6) sutured mandibular symphysis, the posterior edge of which is at the level of the i3-p1 diastema; and 7) ventral border of the mandibular ramus being slightly concave ventrally.
Mystacodon selenensis also bears four additional derived characters: upturned (i.e., dorsally concave) anterior region of the rostrum; posteriormost upper tooth anterior to level of antorbital process of maxilla; broad-based rostrum (ratio between width of skull at rostrum base and width at postorbital process> 0.8); and strong tuberosity on anterior edge of radius (the latter condition is unknown in the other toothed mysticetes).
Furthermore, Mystacodon selenensis bears a series of plesiomophic features: supraoccipital shield not extending anterior to anterior level of squamosal fossa; only two dorsal infraorbital foramina; a basilosaurid-like dental formula 3.1.4.2/3.1.4.3; no wide diastemata between posterior cheek teeth; sutured mandibular symphysis; and well-defined acetabulum on innominate.
Mystacodon selenensis differs from Llanocetus denticrenatus in the following characters: 1) much smaller size (less than half the estimated body size); 2) rostrum narrower with concave edges while the preserved posterior part of the rostrum of Llanocetus is much wider and laterally expanded; 3) alveolar border of the maxilla on posterior region of the rostrum rounded dorsolaterally in cross-section with a subvertical lateral wall, while it is flat and crest-like in Llanocetus; 4) preserved parts of the palate lack deep lateral sulci, while they are distinctly present and abundant in Llanocetus; 5) premaxillae abruptly depressed just anterior to the nasals, while in Llanocetus the transition to the post narial part of the premaxillae is smooth; 6) upper molars closely approximated, while they are separated by large diastemata in Llanocetus; 7) upper molars much larger; 8) short diastema between P3 and P4 (less than one tooth mesiodistal length), while in Llanocetus the diastema is two to three times a tooth length; 9) much longer orbit; 10) nearly straight lateral edge of the supraorbital process, while it is deeply concave in Llanocetus; 11) posterior edge of the supraorbital process roughly straight, while it is concave in Llanocetus; 12) orbitotemporal crests extend posteriorly until anteroposterior mid-point of the parietals, while in Llanocetus they end anteriorly, at the posteromedial angle of the supraorbital process of the frontal; 13) dorsal edge of the parietals is subhorizontal in lateral view, while it ascends steeply toward the vertex in Llanocetus; 14) intertemporal bridge much narrower than long transversely, while it is as wide as long in Llanocetus; 15) anterior angle of nuchal crest posterior to anterior margin of squamosal fossa, while it approximately reaches the level of the middle of the fossae in Llanocetus; 16) lateral edges of the supraoccipital in dorsal view straight, while they are sigmoid in Llanocetus; 17) vertex and dorsal aspect of the braincase almost in the same plane as the posterior region of the rostrum, while in Llanocetus they slope anteriorly and distinctly overhang the rostrum; 18) distal extremity of anterior ribs expanded and pestle-like, while this condition is absent in Llanocetus.
Mystacodon selenensis differs from Coronodon havensteini in having a longer rostrum and longer diastemata between the premolars; much longer nasals associated to more anterior bony nares, located in the anterior half of the rostrum (in the posterior third in Coronodon); temporal fossae more elongated anteroposteriorly; a long and thin (dorsoventrally) zygomatic process of the squamosal (short and stout in Coronodon); a sharp external occipital crest; a tightly articulated mandibular symphysis; and an sub-horizontal or gently sloping tooth wear on all preserved teeth with strong edges at the apical surface.
In addition to its autapomorphies, Mystacodon selenensis differs from mammalodontids in having: a rostrum at least proportionally twice longer; much more anteriorly placed bony nares; a sharply triangular occipital shield; a well-developed external occipital crest (low, when present, in mammalodontids); a strong palatine sulcus on the medial edge of the premaxilla and maxilla anteriorly; much longer diastemata between the premolars; and absence of the markedly V-shaped fronto-parietal suture (as observed in mammalodontids).
As compared with aetiocetids, Mystacodon selenensis has a basilosaurid-like dental formula; an almost horizontal apical tooth wear; much larger postcanine teeth; and a long and slender zygomatic process of the squamosal, which is lowest at its anteriormost portion. M. selenensis lacks all the aetiocetid characteristic features, such as the extreme dorsoventral compression of the rostrum; the deeply concave lateral edge of the supraorbital process of the frontal in dorsal view; the anteroposteriorly short zygomatic process of the squamosal (as compared to that of basilosaurids); and the very thin jugal.
Otuma Formation
Yumaque Member of the Paracas Formation
Los Choros Member of the Paracas Formation
Finally, Mystacodon selenensis lacks cranial synapomorphies of Odontoceti: facial concavity; presence of premaxillary foramen and premaxillary sac fossa; and wide posterior expansion of maxilla overlapping the supraorbital region.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.