
Echinoderes goku, Rucci & Neuhaus & Bulnes, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5099.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:3BD6F566-9B77-48C6-8538-DD3E9BCA08BF |
|
DOI |
https://doi.org/10.5281/zenodo.6309107 |
|
persistent identifier |
https://treatment.plazi.org/id/03C1B533-FFE7-7634-7F8D-FC0DFC679672 |
|
treatment provided by |
Plazi |
|
scientific name |
Echinoderes goku |
| status |
sp. nov. |
Echinoderes goku View in CoL sp. nov.
http://zoobank.org/ urn:lsid:zoobank.org:pub:93EF677E-1755-41C7-8906-952614E71018
Diagnosis. Echinoderes with middorsal spine on segments 4, 6, and 8, and spine in lateroventral positions on segments 6–9 and 11 (lateral terminal spines). Tube present in subdorsal position on segment 2, sublateral to midlateral position on segments 7 and 9, lateral accessory position on segment 8, lateroventral position on segment 5 and ventrolateral position on segment 2. Type-2 gland cell outlets, female-specific papillae or subcuticular funnel-like structures not present. Male with a sublateral tube on segment 10 with a basal cylindrical reinforcement structure and three pairs of penile spines on segment 11. Female with sublateral fringe on surface connected with a subcuticular conical structure surrounded basally by a cylindrical reinforcement structure in segment 10 and with a lateral terminal accessory spine on segment 11.
Etymology. The specific epithet is a noun in apposition and alludes to the adoptive name of the Saiyan Son Gokū, the major character from the Dragon Ball manga, authored by Akira Toriyama.
Material examined. Holotype: Male (MACN-In 43755), collected on March 31, 2019 by Agustín G. Menechella ( IADO, CONICET, Argentina), at station E29 during the campaign aboard the Motorsailer “Bernardo Houssay” through the Argentinean continental shelf ( Fig. 1 View FIGURE 1 ), at 40 m depth using a dredge; mounted as glycerol-paraffin slide on Cobb aluminum frame . Paratypes mounted as glycerol-paraffin slides on Cobb aluminum frames: 1) material deposited at the Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina: one male and two females (catalogue numbers MACN-In 43756a –43756c); 2) material deposited at the Museum für Naturkunde Berlin, Germany, collection “Vermes”, “Generalkatalog freilebende Würmer”: 12 females and 19 males (catalogue numbers ZMB 12262, 12264 View Materials , 12265 View Materials , 12314–12331 View Materials , 12334–12344 View Materials ). Non-types, deposited at Museum für Naturkunde Berlin: four males ( ZMB 12263, 12266 View Materials , 12332 View Materials , 12333 View Materials ) and five juveniles ( ZMB 12267–12269 View Materials , 12345– 12347 View Materials ) mounted as glycerol-paraffin slides on Cobb aluminum frames and two adults of undetermined sex ( ZMB 12270, 12351 View Materials ), one male ( ZMB 12350), and four females ( ZMB 12271, 12272 View Materials , 12348 View Materials , 12349 View Materials ) mounted for SEM on two stubs. All paratypes and non-types collected at same locality and date by same person as the holotype .
Type locality. South West Atlantic Ocean , Argentinian continental shelf, off Pehuen-Có and Monte Hermoso, 39°06.7’S, 061°26.5’W, 40 m depth GoogleMaps .
Description. Adults with head, neck, and 11 trunk segments ( Figs 2A,B View FIGURE 2 , 3A,C View FIGURE 3 , 6A View FIGURE 6 ). For a complete overview of measures and dimensions see Table 2 View TABLE 2 . Distribution of cuticular structures, i.e., sensory spots, gland cells, conical structures, spines and tubes, is summarised in Table 3 View TABLE 3 .
Head. Head consisting of a retractable mouth cone and an introvert ( Figs 3A,C View FIGURE 3 , 6A–C View FIGURE 6 ). Mouth cone equipped with 9 outer oral styles, each with two articulating elements and neighbouring styles slightly alternating in length ( Figs 4K View FIGURE 4 , 6B View FIGURE 6 ). Each outer oral style possessing at its base a posterior central fringe of spinose processes and one anterior lateral spinose process on each side ( Fig. 6B View FIGURE 6 ). Posterior spinose processes may be split apically into two processes ( Fig. 6B View FIGURE 6 ). Although two out of three specimens mounted for SEM appeared with the introvert extended, the information on scalid numbers and arrangement is scarce because of particles partially masking characters. Both odd and even-numbered sectors showing seemingly 6 spinoscalid rings ( Fig. 6C View FIGURE 6 ). All spinoscalids composed of a broader sheath-like base with a terminal fringe of spinose processes and an articulating terminal element ending bluntly in the primary spinoscalids and spinose in rings 02–06 ( Fig. 6C View FIGURE 6 ). Terminal element covered with short cuticular hairs. Each primary spinoscalid revealing apically on its base two prominent fringes of spinose processes ( Fig. 6C View FIGURE 6 ). One prominent apical process also found on the basis of ring 03 spinoscalids but not on those of the following rings. Spinoscalid of ring 06 of each second sector appearing to be much smaller than that of its neighbouring sector. Six short and hairy trichoscalids with trichoscalid plates being present, opposite to the neck placids 2, 6, 8, 10, 12 and 16.
Neck. Neck equipped with 16 trapezoidal placids ( Figs 2A,B View FIGURE 2 , 3E View FIGURE 3 , 4D View FIGURE 4 ), wider posteriorly and with rounded edges anteriorly in SEM of at least one specimen revealing placids at full length (ZMB 12270; not shown here). In light microscopy, anterior end of each placid of almost all specimens looking as if bent partly backwards ( Figs 2A,B View FIGURE 2 , 3E View FIGURE 3 , 4D View FIGURE 4 , 5A View FIGURE 5 ), possibly because of the thin cuticle revealed by most specimens (see below). Midventral placid wider than the remaining ones. All placids articulating with the first trunk segment. Trichoscalid plates present on paradorsal (8 and 10), laterodorsal (6 and 12) and ventromedial (2 and 16) placids.
Trunk. Segments 1 and 2 formed by a closed cuticular ring ( Figs 2A,B View FIGURE 2 , 4A,D View FIGURE 4 , 8C View FIGURE 8 ) and the remaining ones by one tergal and two sternal plates ( Figs 2A,B View FIGURE 2 , 3E,F,J View FIGURE 3 , 4A–E,I View FIGURE 4 , 5A–C View FIGURE 5 , 6A View FIGURE 6 , 7 View FIGURE 7 , 8B,D–F View FIGURE 8 ). Midsternal and tergosternal junctions well-developed, however, the latter being less conspicuous and sometime hard to see under the light microscope. Pachycycli present anteriorly on segments 1–11 ( Figs 3E,F,J View FIGURE 3 , 4A–E,I,J View FIGURE 4 , 5A–C View FIGURE 5 ). Cuticular hairs not abundant and filiform ( Figs 3E,F,J View FIGURE 3 , 5A–C View FIGURE 5 , 6A View FIGURE 6 , 7A–E View FIGURE 7 , 8B–F View FIGURE 8 ). The bracteate hairs on segments 2–10 arranged in one or two irregular wavy rows ( Figs 6A View FIGURE 6 , 7A–E View FIGURE 7 , 8C–F View FIGURE 8 ). Hairs of anterior row shorter than those of the posterior one ( Figs 6A View FIGURE 6 , 7A–E View FIGURE 7 , 8C–F View FIGURE 8 ). Well-developed patch of short, stronger cuticular hairs in three rows present paraventrally and centrally on segments 3–6 ( Figs 4D View FIGURE 4 , 8C–E View FIGURE 8 ) and, less developed, also on segment 7 ( Fig. 8E View FIGURE 8 ). Segments 4– 10 with a ventrolateral to paraventral patch of thin cuticular hairs organised in a single or sometimes in two or three rows ( Fig. 8D–F View FIGURE 8 ). Most adult specimens having moulted recently recognizable by differing extent of fusion of button-like structures in cuticular flap at posterior end of each segment (comp. Figs 4D,F–I View FIGURE 4 , 5A View FIGURE 5 , 9B,D,F–H View FIGURE 9 ). Primary pectinate fringe well-developed in segments 2–11 and formed by teeth usually of even length and width within a segment ( Figs 3E,F,J View FIGURE 3 , 4I View FIGURE 4 , 5B,C View FIGURE 5 , 7 View FIGURE 7 , 8B–F View FIGURE 8 ). Fringe appearing shorter on anterior three segments than in remaining segments and almost lacking dorsally on segment 10 ( Fig. 7D,E View FIGURE 7 ), narrower ventrally on segment 10 than on remaining segments ( Fig. 8F View FIGURE 8 ). Secondary pectinate fringe present as a single row of very short cuticular processes dorsally and ventromedially ( Figs 7C View FIGURE 7 , 8F View FIGURE 8 ). Type-1 sensory spots ( Figs 2 View FIGURE 2 , 3E–J View FIGURE 3 , 4A–D,F–I View FIGURE 4 , 5A–C View FIGURE 5 , 7A–E View FIGURE 7 , 8C–F View FIGURE 8 ) and type-1 gland cell outlets ( Figs 2 View FIGURE 2 , 3E,G,H,J View FIGURE 3 , 4A,B,D–G,I View FIGURE 4 , 5A,B View FIGURE 5 ) present. Type-2 gland cell outlets and female-specific papillae or funnel-like or pear-shaped subcuticular structures absent on any segment.
Segment 1 ( Figs 2A,B View FIGURE 2 , 3E View FIGURE 3 , 4A,D View FIGURE 4 , 5A View FIGURE 5 , 6A View FIGURE 6 , 7A View FIGURE 7 , 8C View FIGURE 8 ; Table 3 View TABLE 3 ) lacking spines and tubes. Sensory spot usually with 2 but sometimes with up to 5 pores present centrally in subdorsal, midlateral and ventromedial position, flanked by long cuticular hairs. Type-1 gland cell outlet occurring centrally in middorsal and anteriorly in lateroventral position. Other hairs than those associated with sensory spots not existing on this segment. Primary pectinate fringe as row of very short teeth with a blunt tip, broader than teeth of remaining segments.
Segment 2 ( Figs 2A,B View FIGURE 2 , 3E View FIGURE 3 , 4A,D View FIGURE 4 , 5A,C View FIGURE 5 , 6A View FIGURE 6 , 7A View FIGURE 7 , 8C View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ) with subdorsal and ventrolateral tube. Sensory spot present posteriorly in middorsal and midlateral position and centrally to posteriorly in ventromedial position. Type-1 gland cell outlet occurring anteriorly in middorsal and ventromedial position. Cuticular hairs scarce on this segment, but more evident if associated with the sensory spots. A few additional hairs emerging from cuticular perforation sites.
Segment 3 ( Figs 2A,B View FIGURE 2 , 3E View FIGURE 3 , 4A,D View FIGURE 4 , 5A,C View FIGURE 5 , 6A View FIGURE 6 , 7A View FIGURE 7 , 8C View FIGURE 8 ; Table 3 View TABLE 3 ) without spines and tubes. Sensory spot present posteriorly in subdorsal and sublateral position. Type-1 gland cell outlet occurring anteriorly in middorsal and centrally in ventromedial position. Cuticular hairs more abundant than on the previous segment and organised similarly on the remaining segments.
Segment 4 ( Figs 2A,B View FIGURE 2 , 3E View FIGURE 3 , 4A,D View FIGURE 4 , 5A,C View FIGURE 5 , 6A View FIGURE 6 , 7B View FIGURE 7 , 8D View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ) with a middorsal acicular spine. Sensory spots lacking on this segment. Type-1 gland cell outlet occurring anteriorly in paradorsal and centrally in paraventral to almost ventromedial position.
Segment 5 ( Figs 2A,B View FIGURE 2 , 3E View FIGURE 3 , 4A View FIGURE 4 , 5A,C View FIGURE 5 , 6A View FIGURE 6 , 7B View FIGURE 7 , 8D View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ) with lateroventral tube. Sensory spot present posteriorly in subdorsal, sublateral to midlateral position and almost centrally in ventromedial position. Sublateral to midlateral spot of segments 5 and 6 slightly more dorsally than spot of segment 3. Type-1 gland cell outlets occurring anteriorly in middorsal and centrally in paraventral to almost ventromedial position.
Segment 6 ( Figs 2A,B View FIGURE 2 , 3F,J View FIGURE 3 , 4B View FIGURE 4 , 5B,C View FIGURE 5 , 6A View FIGURE 6 , 7B View FIGURE 7 , 8D View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ) with middorsal and lateroventral acicular spine. Sensory spot present posteriorly in paradorsal and sublateral to midlateral position and almost centrally in ventromedial position. Type-1 gland cell outlet occurring anteriorly in paradorsal and centrally in paraventral to almost ventromedial position.
Segment 7 ( Figs 2A,B View FIGURE 2 , 3F,J View FIGURE 3 , 4B View FIGURE 4 , 5B,C View FIGURE 5 , 6A View FIGURE 6 , 7C View FIGURE 7 , 8D View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ) with sublateral to midlateral tube and lateroventral acicular spine. Sensory spot present posteriorly in subdorsal position and almost centrally in ventromedial position. Type-1 gland cell outlet occurring anteriorly in middorsal and paraventral to almost ventromedial position.
Segment 8 ( Figs 2A,B View FIGURE 2 , 3F,J View FIGURE 3 , 4B,I,J View FIGURE 4 , 5B View FIGURE 5 , 6A View FIGURE 6 , 7C View FIGURE 7 , 8D,E View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ) with a middorsal acicular spine, lateral accessory tube and lateroventral acicular spine. Middorsal spine of female much longer (92–98 µm) than that of male (54–79 µm) ( Table 2 View TABLE 2 ). Sensory spot present posteriorly in paradorsal position. Type-1 gland cell outlet occurring anteriorly in paradorsal and paraventral to almost ventromedial position.
Segment 9 ( Figs 2 View FIGURE 2 , 3F,G,J View FIGURE 3 , 4B,C,I,J View FIGURE 4 , 5B,F,G View FIGURE 5 , 6A View FIGURE 6 , 7D View FIGURE 7 , 8E,F View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ) with sublateral to midlateral tube and lateroventral acicular spine. Sensory spot present posteriorly in paradorsal and centrally in laterodorsal and ventrolateral position. Type-1 gland cell outlet occurring anteriorly in paradorsal and centrally in paraventral position.
Segment 10 ( Figs 2 View FIGURE 2 , 3G,H,J View FIGURE 3 , 4B,C,E–J View FIGURE 4 , 5B,D,E View FIGURE 5 , 6A View FIGURE 6 , 7D–F View FIGURE 7 , 8B,F View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ) without spines but with sublateral to midlateral straight short tube with basal cuticular cylindrical reinforcement structure below trunk cuticle in male ( Fig. 4F,H View FIGURE 4 ). Female without tube but with sublateral to midlateral subcuticular, conical structure ( Figs 5D,E View FIGURE 5 , 10C View FIGURE 10 ) ending in short apical cuticular duct through trunk cuticle, with basal cuticular cylindrical reinforcement structure below trunk cuticle ( Fig. 5D,E View FIGURE 5 ), opening to outside via fringed pore ( Fig. 7E,F View FIGURE 7 ). Sensory spot present posteriorly in subdorsal and ventrolateral position. Type-1 gland cell outlet occurring anteriorly in middorsal and centrally in middorsal and paraventral position. Posterior margin of each sternal plate progressing more posteriorly midventrally than lateroventrally.
Segment 11 ( Figs 2 View FIGURE 2 , 3H,I View FIGURE 3 , 4F–J View FIGURE 4 , 5D,E View FIGURE 5 , 6A View FIGURE 6 , 7D,E View FIGURE 7 , 8A,B,F View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ) with lateral terminal spine, varying considerably in length (146–208 µm) in both females and males ( Table 2 View TABLE 2 ). Straight lateral terminal accessory spine present in females ( Figs 2C,D View FIGURE 2 , 4I,J View FIGURE 4 , 5D View FIGURE 5 , 6A View FIGURE 6 , 7D,E View FIGURE 7 , 8F View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ) and three penile spines appearing in males, one bent, tubular, longer, in lateral accessory position, one shorter, broader, almost triangular, slightly more dorsally, and one bent, tubular, longer sublaterally ( Figs 2A,B View FIGURE 2 , 4F,G View FIGURE 4 , 8A View FIGURE 8 ; Tables 2 View TABLE 2 , 3 View TABLE 3 ). Sensory spot present posteriorly in subdorsal position. Males often with two ventrolateral pores, unclear if belonging to a sensory spot. Two type-1 gland cell outlets occurring anteriorly and centrally behind each other in middorsal position. Tergal extensions triangular-shaped.
Among the 39 specimens studied by LM and three specimens investigatable with SEM no variation of characters was found except that the position of sensory spots on segment 5 and 6 and of tubes on segments 7 and 9 varied slightly from more sublaterally to more midlaterally. Probably, one specimen ( ZMB 12323) possessed a lateroventral spine only on the right side of segment 7 .
Juvenile stages. Five specimens could be identified as juvenile stages based on their thin trunk cuticle, the postmarginal spicula, a gap in the arrangement of cuticular hairs and postmarginal spicula about ventromedially to paraventrally where the dorsoventral muscles attach ( Fig. 9B,C,E View FIGURE 9 ), papillate sensory spots ( Fig. 10A View FIGURE 10 ) if present at all, the lack of recognisable gland cells, an acicular spine middorsally on segment 9 ( Fig. 10A View FIGURE 10 ) and lateral accessorily on segment 10 ( Fig. 10C View FIGURE 10 ), and a well developed lateral terminal accessory spine ( Fig. 9A,E,F View FIGURE 9 ). It was not possible to identify different life history stages with certainty, because the specimens came out crumpled, twisted, and folded during mounting due to their thin cuticle. In all specimens, 11 trunk segments delimited by postmarginal spicula could be recognised, whereas sensory spots were difficult to trace. Four specimens seemed to represent late juvenile stages if not the last stage, and the fifth was moulting to a female, and this is subsequently described in more detail and documented.
Last juvenile stage moulting. One specimen was fixed while moulting from the last juvenile stage to the adult stage (ZMB 12267, Fig.9A View FIGURE 9 ). Each of the 11 trunk segments of the juvenile specimen was covered by numerous short cuticular hairs in its central part and showed a row of postmarginal spicula posteriorly on each segment ( Figs 9B–H View FIGURE 9 , 10A,B View FIGURE 10 ). An acicular spine occurred middorsally on segments 4, 6, 8 and 9 ( Figs 9H View FIGURE 9 , 10A View FIGURE 10 ), in lateral accessory position on segments 10 ( Fig. 10C View FIGURE 10 ) and 11 (= lateral terminal accessory spine), and lateroventrally on segments 6–9 and 11 (= lateral terminal spine) ( Figs 9 View FIGURE 9 , 10A View FIGURE 10 ). A straight tube was found sublaterally on segments 7, 9 and 10 ( Fig. 10B View FIGURE 10 ), lateral accessorily on segment 8, lateroventrally on segment 5, and ventrolaterally on segment 2. A papillate sensory spot appeared at least paradorsally on segments 6, 8 and 9 ( Fig. 10A View FIGURE 10 ), subdorsally on segments 10 and 11 and possibly on segment 5, ventrolaterally on segments 9–11, and ventromedially on segments 1 and 5–7.
The moulting adult possessed in the free flap of segments 1–10 the button-like structures, which are typical of recently moulted adults ( Fig. 9B,D,G,H View FIGURE 9 ). The adult lacked the middorsal spine on segment 9 and the lateral accessory spine on segment 10 as well as the sublateral tube on segment 10. In the latter position, it revealed a subcuticular conical structure with a basal cuticular reinforcement structure ( Fig. 10C View FIGURE 10 ).
Additional material. A reinvestigation of sublateral to midlateral subcuticular characters in segment 10 of 14 species of Echinoderidae revealed the following results: (1) Female and male Echinoderes unispinosus unveiled a subcuticular conical structure ( Fig. 5J View FIGURE 5 ). (2) Female and male E. meteorensis had a subcuticular very small conical structure plus a cylindrical reinforcement structure at the base of the laterodorsal tube ( Fig. 5H,I View FIGURE 5 ). (3) Female and male E. apex possessed a subcuticular spherical structure at the base of the laterodorsal tube ( Fig. 5K,L View FIGURE 5 ). (4) Female and male E. capitatus and E. ferrugineus expressed a subcuticular cylindrical reinforcement structure at the base of the laterodorsal tube ( E. capitatus , Fig. 5F,G View FIGURE 5 ) or a tube was missing ( E. ferrugineus , Fig. 5M View FIGURE 5 ). (5) Female and male E. pilosus exhibited a subcuticular reinforcement structure at the base of the laterodorsal tube plus an external cylindrical cuticular sheath surrounding the basal part of the tube ( Fig. 5N View FIGURE 5 ). (6) Female Cephalorhyncha asiatica , Echinoderes applicitus , E. bathyalis , E. multiporus , E. pterus , Fissuroderes higgins , F. thermoi , and Polacanthoderes martinezi did not show any subcuticular structures, males were not studied.
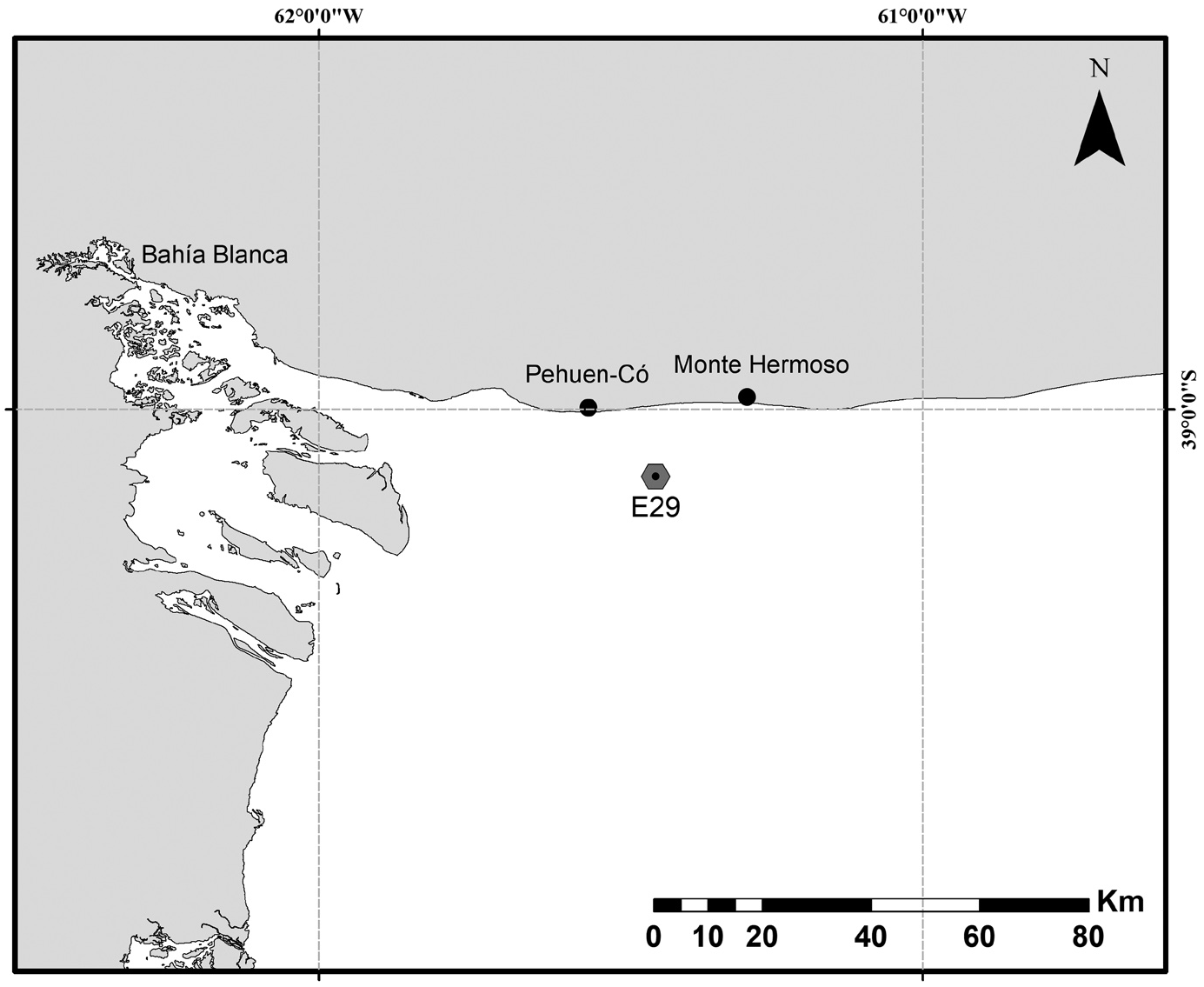
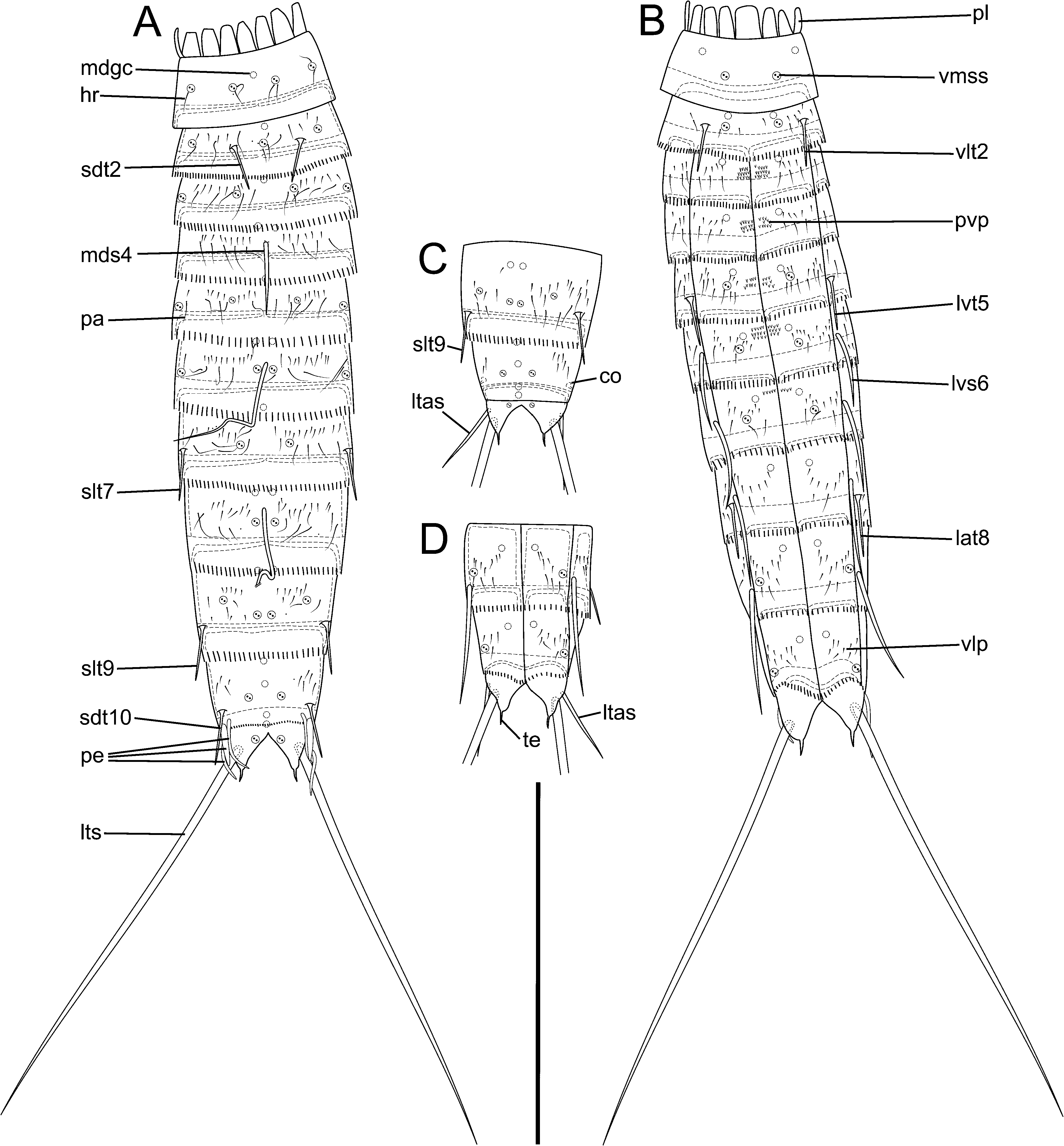



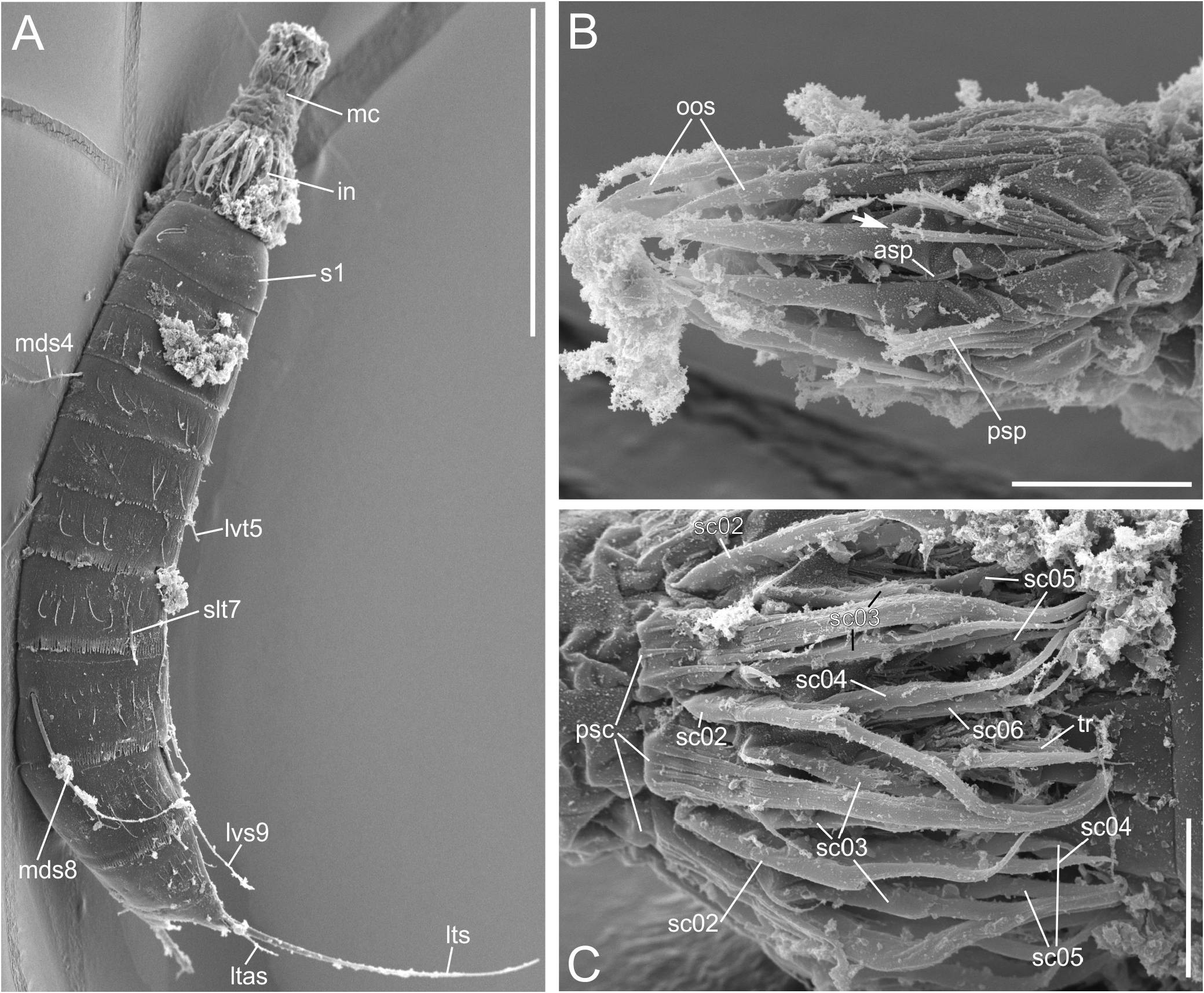
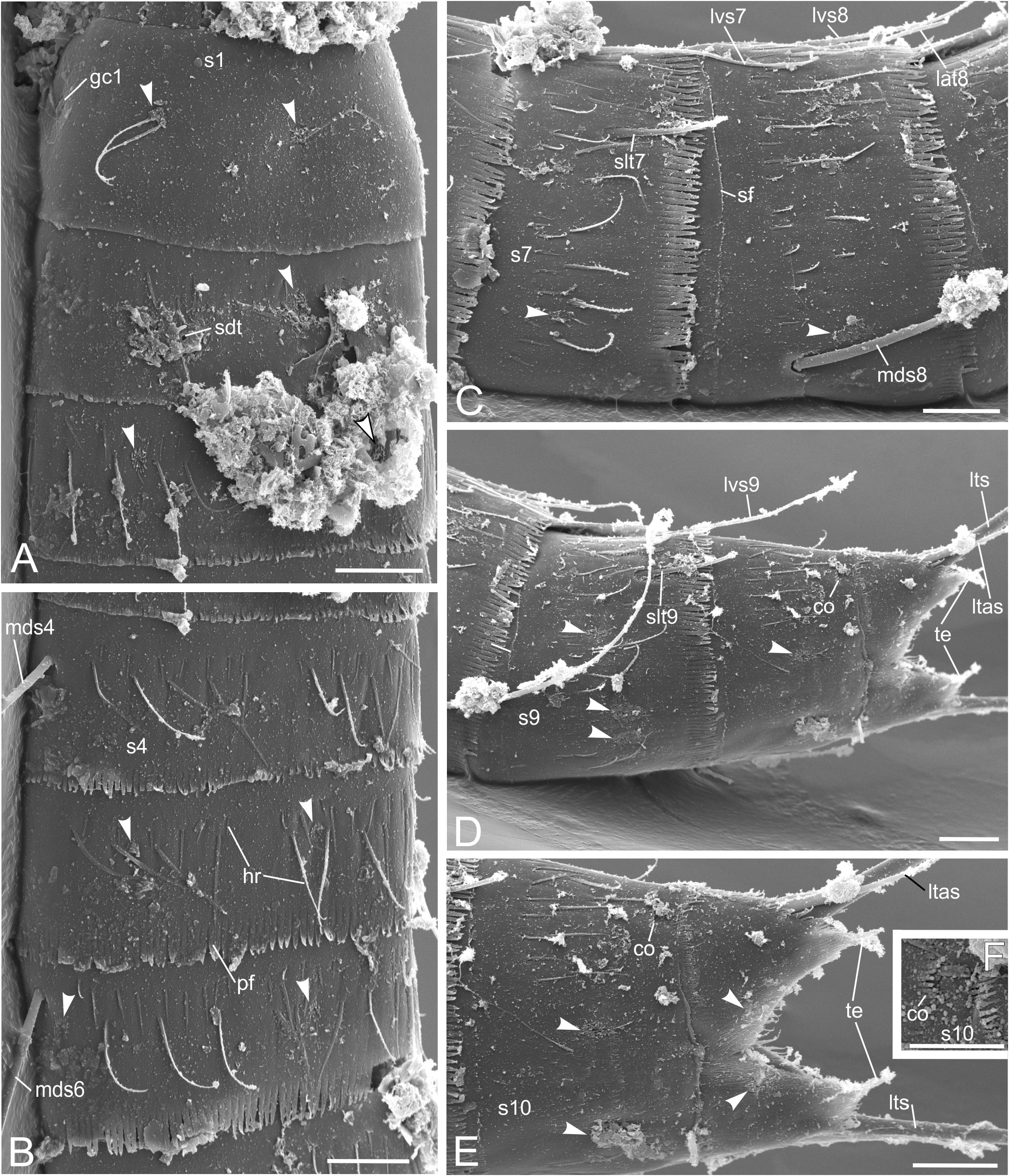



TABLE 3. Summary of nature and location of sensory spots, gland cell outlets and spines arranged by series in Echinoderes goku sp. nov. from Argentina. Features appearing anteriorly on a segment do not show any background, features located centrally on a segment are indicated by a grey background, and characters occurring posteriorly on a segment are marked in black. Sensory spots on segments 5 and 6 and tubes on segments 7 and 9 are located in a sublateral to almost midlateral position. Abbreviations: ♀ or ♂, female or male condition of sexually dimorphic character; ac, acicular spine; co, fringe on trunk surface with conical subcuticular structure and basal cylindrical reinforcement structure in female; gc1, type-1 gland cell outlet; la, lateral accessory; ld, laterodorsal; ltas, lateral terminal accessory spine; lts, lateral terminal spine; lv, lateroventral; md, middorsal; pd, paradorsal; pe, penile spine; pv, paraventral; sd, subdorsal; sl, sublateral; ssp1, type-1 sensory spot; tu, tube; tuc, tube with basal cylindrical reinforcement structure in male; vl, ventrolateral; vm, ventromedial.
| Position Segment | md | pd | sd | ld | ml | sl | la | lv | vl | vm | pv |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | gc1 | ssp1 | ssp1 | gc1 | ssp1 | ||||||
| 2 | gc1; ssp1 | tu | ssp1 | tu | ssp1; gc1 | ||||||
| 3 | gc1 | ssp1 | ssp1 | gc1 | |||||||
| 4 | ac | gc1 | gc1 | ||||||||
| 5 | gc1 | ssp1 | ssp1 | tu | ssp1 | gc1 | |||||
| 6 | ac | gc1; ssp1 | ssp1 | ac | ssp1 | gc1 | |||||
| 7 | gc1 | ssp1 | tu | ac | ssp1 | gc1 | |||||
| 8 | ac | gc1; ssp1 | tu | ac | gc1 | ||||||
| 9 | gc1; ssp1 | ssp1 | tu | ac | ssp1 | gc1 | |||||
| 10 | gc1; gc1 | ssp1 | ♂: tuc; ♀: co | ssp1 | gc1 | ||||||
| 11 | gc1; gc1 | ssp1 | ♀: ltas; ♂: 3x pe | lts |
| ZMB |
Museum für Naturkunde Berlin (Zoological Collections) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |