
Bohra sp.
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5299.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:9CA85AEC-7128-4118-A50D-FCD16502F5E0 |
|
DOI |
https://doi.org/10.5281/zenodo.8017949 |
|
persistent identifier |
https://treatment.plazi.org/id/03C24E22-F65B-5641-FF01-C5BBC5D9F276 |
|
treatment provided by |
Plazi |
|
scientific name |
Bohra sp. |
| status |
|
Bohra sp. indet. 2
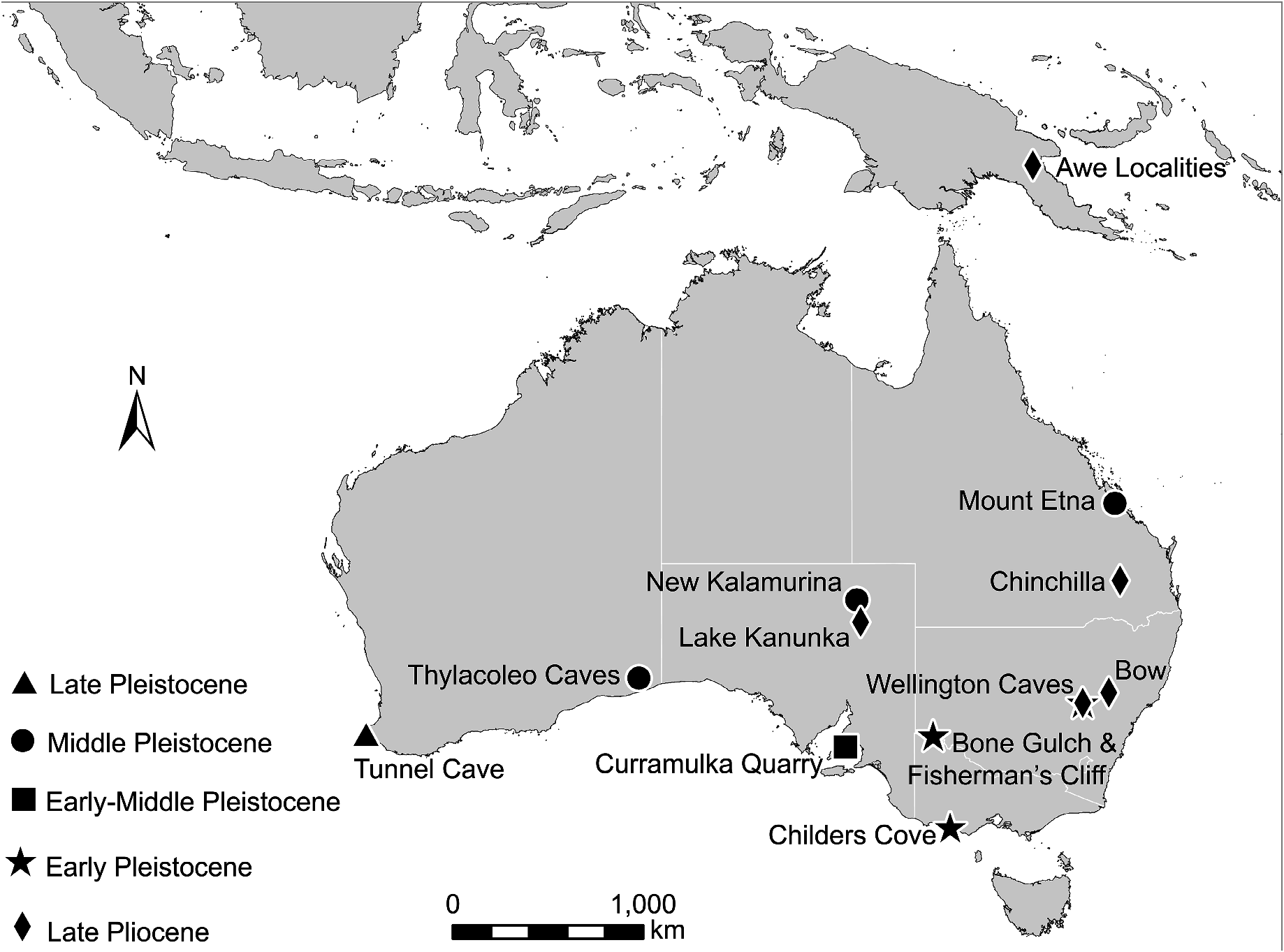
Referred specimen. UCMP 60850 View Materials a, right talus . SAM Hill (= Lake Kanunka Site 2, UCMP V5773 ), Lake Kanunka , eastern Lake Eyre Basin , South Australia ( Figure 1 View FIGURE 1 ). This specimen was collected by Richard H. Tedford et al. on 16 August 1961 .
Description and comparisons. In overall morphology, the talus is dorsoventrally compressed (especially in cranial view) with low trochlear crests, subequal in height and very shallow trochlea groove ( Figure 35 View FIGURE 35 ). The malleolar fossa is large, longer than wide, and moderately deep. The medial malleolus is broad and flattened. The neck of talus is intermediate in length, and extends cranially rather than craniomedially, such that lateral margin of navicular facet overlaps with medial trochlear crest from cranial view. The navicular facet of talar head is obliquely aligned relative to the craniocaudal plane, and relatively narrow and elongate. The talocalcaneal facets are smoothly conjoined and relatively shallow. The posterior plantar process is blunt (short) and broad.
UCMP 60850a is intermediate between the holotype tali of B. illuminata and B. nullarbora in size and morphology, differing from the former by having a wider malleolar fossa and from the latter by bearing confluent calcaneal facets. The trochlear groove is more shallow than, but not as wide as, in B. illuminata , B. nullarbora , D. bennettianus and D. lumholtzi , but similar to B. planei . The medial trochlear crest is lower than in B. paulae , and is more similar to B. planei . The wide malleolar fossa is similar to D. bennettianus . The medial malleolus is relatively less projecting than in B. nullarbora , Dendrolagus , Dorcopsulus , Setonix , Thylogale , but similar to B. paulae and B. planei . The neck of talus is longer than in other species of Bohra and Dendrolagus , and the navicular facet of the talar head is narrower and more elongate, and less obliquely orientated than in B. paulae and B. planei , and particular B. illuminata and B. nullarbora . Although slightly water worn and abraded, the tibiotalar ligament fossa on the medial face of UCMP 60850a appears to have been very shallow and not extended above the dorsal border of the malleolus. The form of this fossa is most similar to the condition observed in B. planei in its medial restriction, but UCMP 60850a differs by lacking a distinct dorsal margin.
Remarks. UCMP 60850a has suffered some post-depositional abrasion due to the fluviatile nature of the deposit, but it is complete enough to confidently say that it belongs to a species of Bohra . Although its morphology is not consistent with the talus of any of the four species for which this element is known ( B. paulae , B. illuminata , B. nullarbora , B. planei ), it is too incomplete to put a species name on. Nevertheless, UCMP60850a represents a definitive record for the genus in the late Pliocene of inland Australia. This is significant, because specimens from the Tirari Formation of the Lake Eyre Basin previously identified as belonging to large tree-kangaroos can now be attributed to other taxa. A maxilla fragment from Stirton Quarry (AMNH 144684 = SIAM 70) referred to cf. Dendrolagus ( Tedford et al. 1992) belongs to a sthenurine (GJP, pers. obs.), while metatarsals from the Toolapinna LF tentatively referred to Dendrolagus ( Tedford et al. 1992) belong to a species of Protemnodon (A. B. Camens, pers. comm., 2019; NMW, pers. obs.).
Phylogenetic analysis
Analysis of the 42 species and 85 characters (Appendices 1–2) produced 168 equally most parsimonious trees with a length of 251 steps, consistency index of 0.56, and retention index of 0.83. The strict consensus tree ( Figure 36 View FIGURE 36 ) collapses multiple nodes, the majority of which are within the Dendrolagina . Bohra is problematic due to the large proportion of missing data for most species, whereas Dendrolagus is challenging due to the markedly uniform osteology of its species.
Macropodinae is supported by four unique synapomorphies ( Figure 36 View FIGURE 36 ): I3 bears a distinct anterobuccal crest (character 15); i1 rests on the palate, bordered by I2–3, during cheek-tooth occlusion (character 38); the ischium and ilium are aligned in the same plane (character 77); the ulnar olecranon process is moderately proportioned (character 85). Macropodines are also united by the possession of either a slightly or markedly broadened I1 (character 14), and by shared possession of bilophodont molars that lack a urocrista, a condition convergently characteristic of the sthenurine Hadronomas puckridgi Kear, 2002 (character 25). Macropodinae is supported by a bootstrap value of 72.
Dorcopsoides fossilis Woodburne, 1967 is united with a Dorcopsis – Dorcopsulus – Watutia clade ( Dorcopsini ) as the basal macropodine branch in 64% of trees, where they are united by two unique synapomorphies: lateral constriction of P3 immediately anterior to the posterolingual cusp (character 21); a deep masseter muscle that extends to below the posterior cheek teeth, with the masseteric canal posteriorly confluent with the dental canal (character 35). In the strict consensus tree, D. fossilis forms a trichotomy with the dorcopsins and a clade giving rise to all other macropodines ( Figure 36 View FIGURE 36 ). Dorcopsis , Dorcopsulus and Watutia are united by two unique synapomorphies: the main crest of P3 is characterised by four cuspules with coarse ridgelets that extend to the crown base (character 17); the P3 crown is markedly deeper anteriorly than posteriorly (character 19).
The clade giving rise to all other macropodines is supported by one unique synapomorphy (I1 markedly broadened; character 14) and four homoplastic features ( Figure 36 View FIGURE 36 ). Setonix brachyurus is reconstructed as the sister taxon to Dendrolagini + Macropodini in 82% of trees, but forms a trichotomy with those two tribes in the strict consensus tree. The monophyly of a clade composed of Wallabia Trouessart, 1904 , Lagorchestes Gould, 1841 , Prionotemnus Stirton, 1955 , Kurrabi Flannery & Archer, 1984 , Baringa Flannery & Hann, 1984 , Onychogalea Gould, 1841 , Osphranter and Macropus (Macropodini) is solidly supported (bootstrap value 74) by three unique podial synapomorphies: relatively short articular region for the talus on the calcaneus (character 53); rounded sustentaculum tali (character 60); medial malleolus on the talus characterised by a narrow, deep fossa and a small process (character 65) ( Figure 36 View FIGURE 36 ).
Thylogale , Petrogale , Dendrolagus and Bohra (Dendrolagini) are united by two unique synapomorphies: triangular inflation of the squamosal between the occiput and zygomatic arch (character 11); relatively broad calcaneocuboid articulation (character 56). This clade is not supported in the bootstrap analysis.
Dendrolagus and Bohra (Dendrolagina) are united by three unique synapomorphies ( Figure 36 View FIGURE 36 ), each a feature of the calcaneus. These are: posteromedial expansion of the tuber calcanei (character 51); relatively broad calcaneus–talus articulation (character 57); marked medial taper of the lateral talar facet (character 59). This alliance is reinforced by several homoplastic synapomorphies ( Figure 36 View FIGURE 36 ) and supported by a bootstrap value of 70. Bohra is recovered as paraphyletic in all 168 trees, and all species of Bohra collapse to form a comb with Dendrolagus in the bootstrap reconstruction due to having support values <50%. However, in the strict consensus tree, B. illuminata , B. nullarbora and B. paulae form a clade with Dendrolagus , united by three unique synapomorphies: smooth calcaneocuboid articulation (character 54); ventromedial facet of cuboid articulation on calcaneus reduced and confluent with dorsomedial facet (character 55); talus with medial malleolar fossa broad and moderately deep with large malleolar process (character 65).
The species of Dendrolagus are united by four unique synapomorphies ( Figure 36 View FIGURE 36 ): a deep cleft on P3/p3 immediately posterior to the anterior cusp of the main crest (character 18); a relatively broad metatarsal V (character 69); a relatively long anterior crest on tibia (character 72); a short tibia relative to the femur (character 74). Two homoplastic traits may also be considered solid synapomorphies for the clade: a relatively shallow sustentaculum tali on the calcaneus otherwise only characterises the outgroup Hypsiprymnodon moschatus (character 62), while a short metatarsal IV relative to the length of the calcaneus is only seen elsewhere in Setonix brachyurus (character 66). Within Dendrolagus , the strict consensus tree produces a trichotomy between D. inustus , a D. bennettianus – D. lumholtzi clade and a clade composed of D. matschiei F̂rster & Rothschild, 1907, D. dorianus Ramsay, 1883 , D. goodfellowi Thomas, 1908 , D. scottae Flannery & Seri, 1990 and D. mbaiso . The New Guinea clade (excluding D. inustus ) is united by two solid synapomorphies: relatively short distal fibular facet on the tibia (character 71); short pes (character 76). The latter is homoplastic, but only with the evolutionarily distant outgroup Hypsiprymnodon moschatus , and so is a very reliable synapomorphy.
TABLE 9. (Continued)
TABLE 9. (Continued)
Body-size estimation
Calcaneal measurements and body mass are highly positively correlated (p <0.05) within extant tree-kangaroos and macropodines as a whole ( Table 10 View TABLE 10 ). This engenders confidence in their utility for modelling body masses of fossil kangaroos. The correlation between femur circumference and body mass is weaker than expected (r 2 = 0.31) within tree-kangaroos and is not significant (p = 0.06), although it is significant within the larger macropodine sample (r 2 = 0.91). Ratio estimators range from 0.9649 to 1.4487 ( Table 10 View TABLE 10 ). Corrected body-mass predictions vary depending on the dataset used to generate them, with those estimated from the modern tree-kangaroo dataset larger than those estimated from the macropodine dataset ( Table 11 View TABLE 11 ). Body-mass estimates predicted from measurements where r 2 ≥ 0.70 fall within the following ranges: 25.7–43.5 kg for Bohra paulae ; 25.6–47.4 kg for B. wilkinsonorum ; 16.0– 26.9 kg for B. nullarbora ; 15.9–26.7 kg for B. planei ; and 18.7–29.9 kg for B. sp. indet. 1 ( Table 11 View TABLE 11 ). No body-mass estimates were generated for B. illuminata , because the holotype is a young adult with unfused epiphyses, including that of the calcaneus.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.