
HAMINOEIDAE Pilsbry, 1895a
|
publication ID |
https://doi.org/ 10.1080/00222933.2018.1533598 |
|
publication LSID |
lsid:zoobank.org:pub:90C191A9-5CA6-4BC6-84E3-B1A26674FA1B |
|
persistent identifier |
https://treatment.plazi.org/id/03C2878D-5C22-152C-FE7F-FA540580BC6A |
|
treatment provided by |
Felipe |
|
scientific name |
HAMINOEIDAE Pilsbry, 1895a |
| status |
|
Family HAMINOEIDAE Pilsbry, 1895a View in CoL
Genus Haminoea Turton and Kingston [in Carrington], 1830
Haminoea aptei sp. nov.
( Figures 1 – 3 View Figure 1 View Figure 2 View Figure 3 )
Diagnosis
Animal light olive with dark green blotches; mantle with brown, green and black dots, and fine white spots; cephalic deeply bilobed; visible eyes, unpigmented periocular area; Hancock ’ s organ, simple horizontal ridge. Shell pale yellowish, translucent; periostracum light orange to cream coloured; columellar lip thin, slightly reflected; columellar callus broad; outer lip, rounded, extends beyond apex. Jaws crescent shaped, formed by rodlet-like structures; ends of rodlets diamond shaped, with small indentations. Radular formula 18 × 6.1.1.1.6; rachidian tooth with three triangular cusps, larger central cusp; lateral teeth hook-shaped, smooth, tapering outwardly; inner lateral with broader cusp. Gizzard plate with 19 ridges; rachis present. Both upper sides of ridges and rachis covered by pointed rods; rods slightly sparsely distributed on top of ridges; absent on lower part of ridges. Male reproductive system with thick-walled atrium, thin-walled fundus; lower fundus narrow, upper fundus oval to bulbous; fundus filled with soft warts, forming vertical ridges, right side of upper fundus with larger, slightly pointed warts, separated by thick horizontal ridges and thin vertical ridges.
Type locality
Burmanallah , South Andaman, Andaman Islands, India (11.574511°N, 92.738906°E) GoogleMaps .
Etymology
The species is named after Dr Deepak Arun Apte, current Director of the Bombay Natural History Society, India, for his excellent and dedicated work on the taxonomy of Indian Mollusca including opisthobranchs.
Material examined
India, Burmanallah , Andaman and Nicobar Islands (11.574511°N, 92.738906°E), 1 spc GoogleMaps ., BNHS opistho 1343A (holotype), H = 6 mm, 24 March 2016; 26 spcs (6 dissected), BNHS opistho 1343/WII/NWR/ BNHS _9 (paratypes), H = 7.1 – 7.9 mm, 24 March 2016; 7 spcs (2 dissected, 2 sequenced), BNHS opistho 1344/WII/NWR/ BNHS _31, H = 6.0 – 8.2 mm, 8 March 2016 . India, Lakshadweep, Minicoy Island (8.27333333°N, 73.02638889°E), 3 spcs and 1 shell (2 dissected and sequenced), BNHS opistho 1361, H = 8 – 13 mm, 18 September 2014 GoogleMaps .
COI barcodes
GenBank accession numbers MH 638603 View Materials ( BNHS opistho 1344), MH 638604 View Materials ( BNHS opistho 1344), MH 638590 View Materials ( BNHS opistho 1361), MH 638591 View Materials ( BNHS opistho 1361).
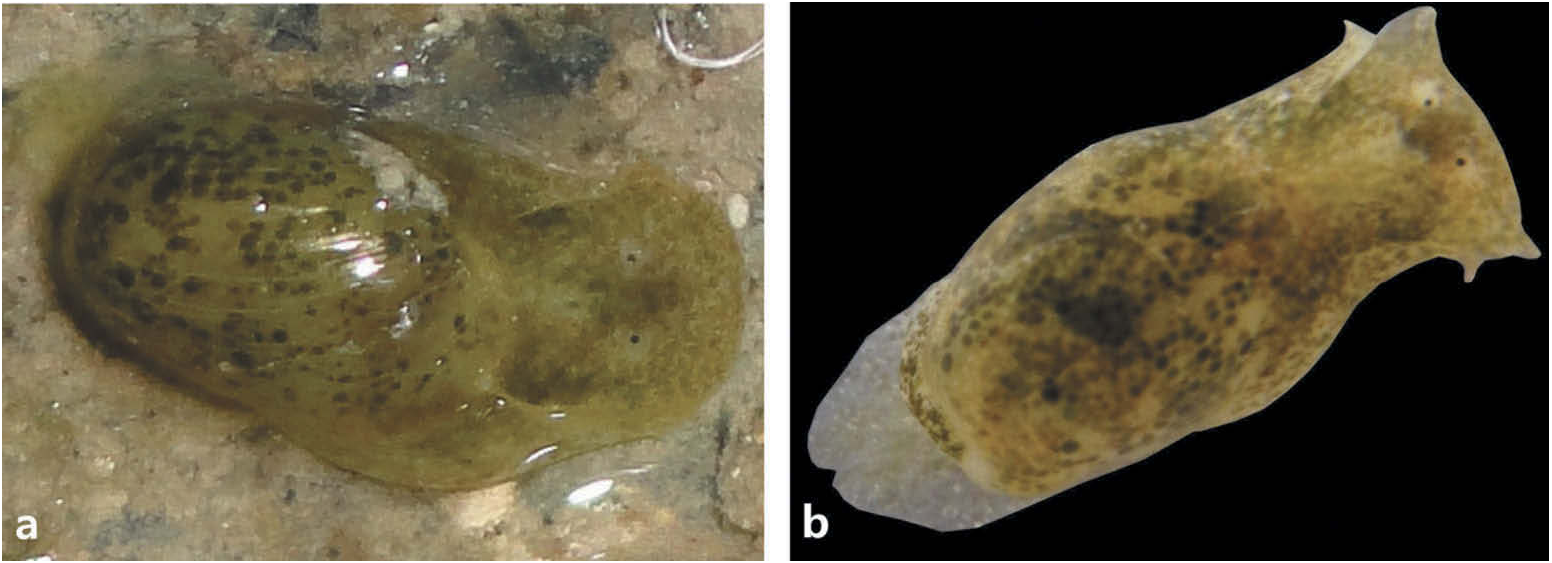
External morphology ( Figure 1 View Figure 1 )
Animal light olive with dark green blotches; mantle with brown, green, black dots, and fine white spots. Cephalic shield broad anteriorly, laterally funnel-like structures present in some specimens only; tapering posteriorly, deeply bilobed, with cephalic lobes extending over anterior part of shell. Eyes visible, unpigmented periocular area.
Hancock ’ s organ simple, horizontal ridge. Parapodial lobes do not meet dorsally. Rounded pallial lobe, extends beyond apex.
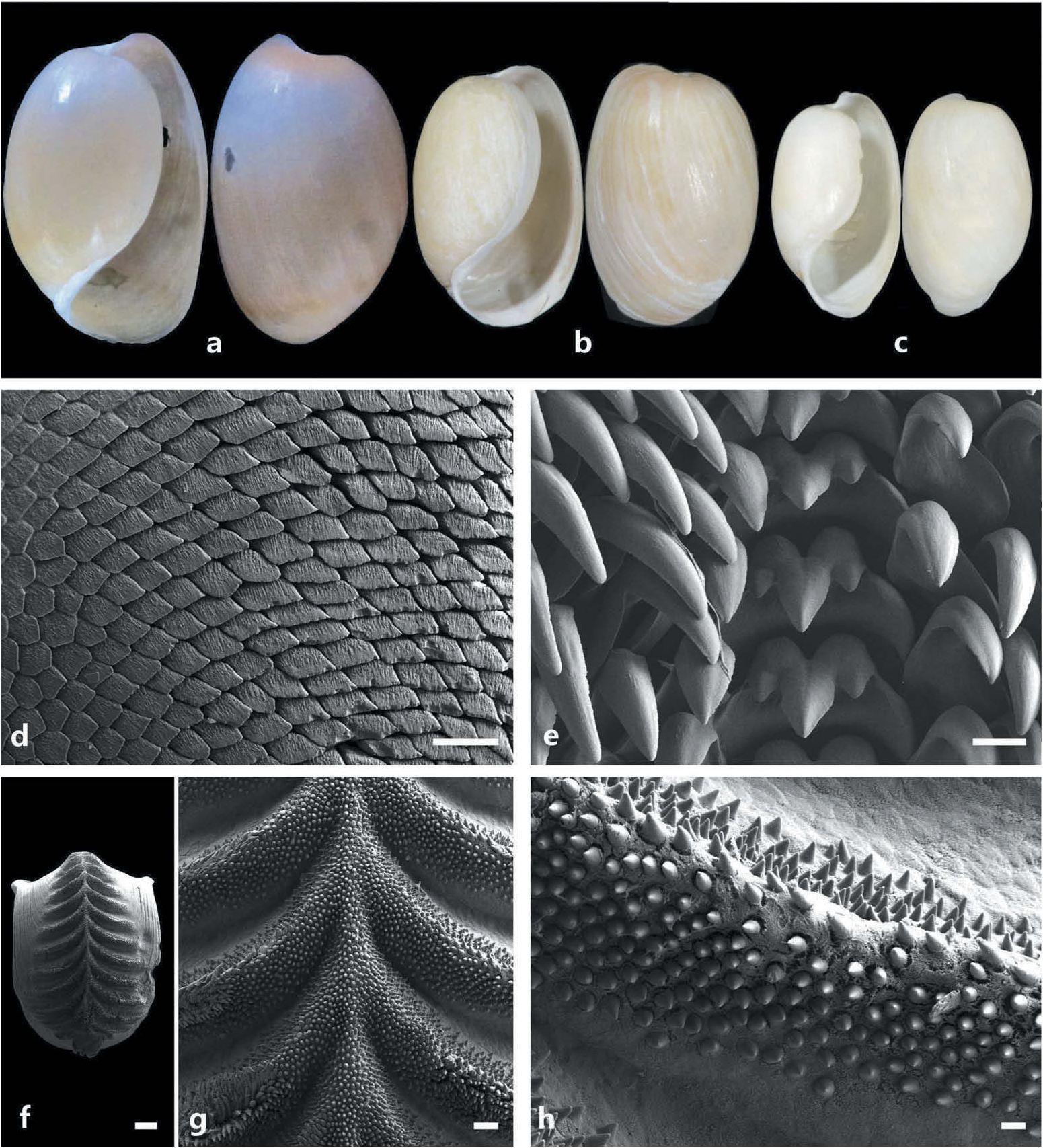
Shell ( Figure 2 View Figure 2 (a–c))
Shell, pale yellowish, translucent; periostracum light orange to cream coloured; shape cylindrical-elongate in smaller specimens (H = 6 mm) to oval in larger specimens (H = 8 – 13 mm). Aperture, broad anteriorly, tapers posteriorly; columellar lip thin, slightly reflected; columellar callus broad; outer lip rounded, extends beyond apex; shoulder rounded.
Jaws ( Figure 2 View Figure 2 (d))
Crescent shaped, formed by rodlet-like structures; ends of rodlets diamond shaped, with small indentations.
Radula ( Figure 2 View Figure 2 (e))
Radular formula 18 – 26 × 6 – 5.1.1.1.5 – 6 (based on three specimens with shell H = 7.9, 7.9, 8.2 mm). Rachidian tooth with three triangular cusps, central cusp larger. Lateral teeth hook shaped, smooth; inner lateral with broader cusp; outer laterals slimmer, tapering outwardly.
Gizzard plates ( Figure 2 View Figure 2 (f–h))
Plates with 19 ridges. Rachis present. Both upper sides of ridges and rachis covered by pointed rods; rods slightly more sparsely distributed on top of ridges; absent on lower part of ridges.
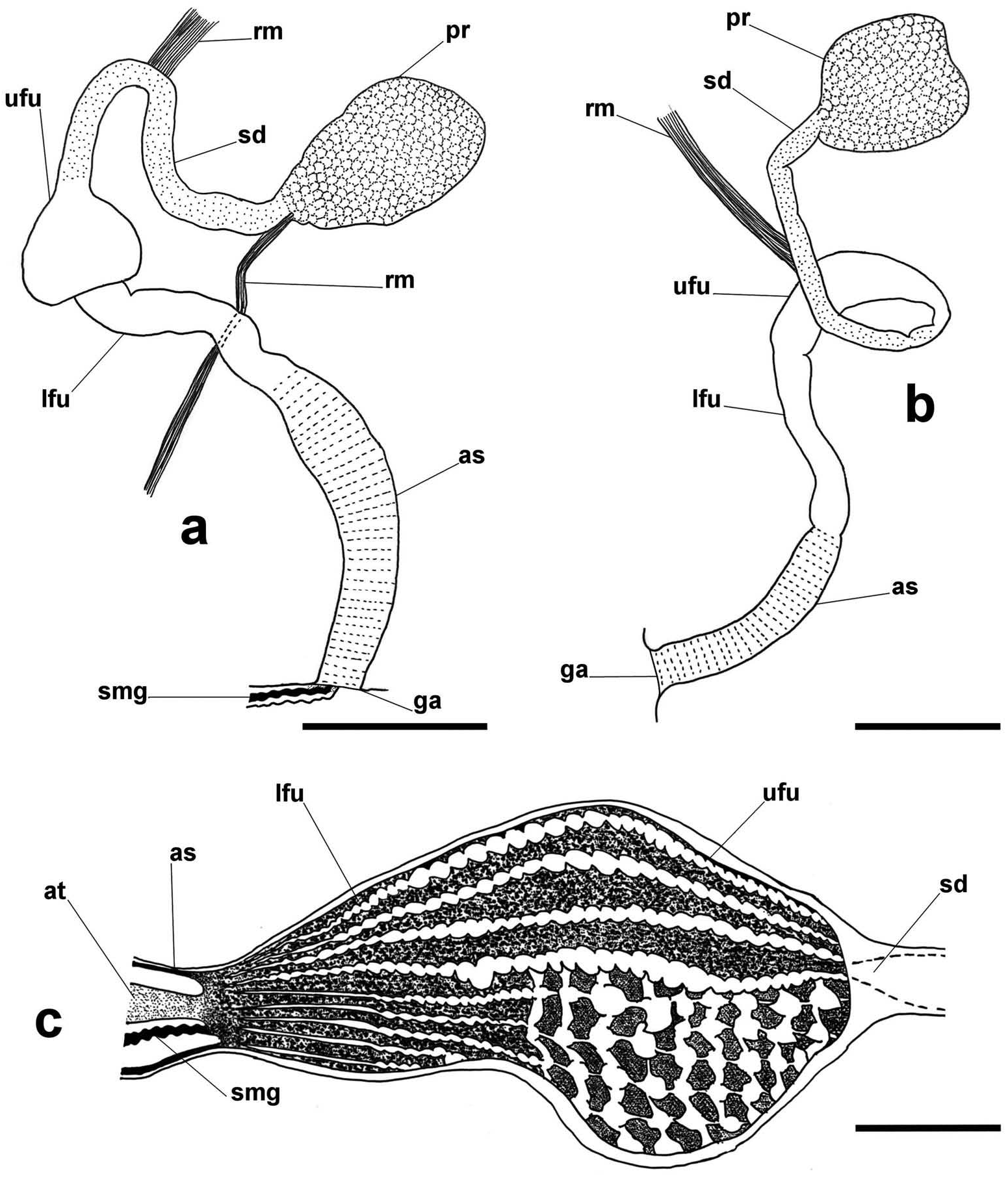
Male reproductive system ( Figure 3 View Figure 3 (a–c))
Consists of atrium, fundus, seminal duct and prostate. Atrium with thickened walls. Fundus with lower and upper parts, both thin walled; lower fundus narrow; upper fundus broader with variable shape, oval-elongated to bulbous; fundus filled with soft warts, forming axial ridges; warts tapers in size towards atrium; right side of upper fundus with larger, slightly pointed warts, separated by thick transversal ridges and thin axial ridges. Two retractor muscles; one connects seminal duct to mantle, the other to proximal region of atrium sheath.
Ecology
Rocky intertidal shores with cyanobacteria mats, green and red algae.
Distribution
So far only known from India in the Andaman and Nicobar Islands (Bay of Bengal) and Minicoy Island, Lakshadweep Islands ( Laccadive Sea).
Phylogenetic and species delimitation analyses
Both COI analyses (with and without third codon positions included) resulted in similar tree topologies, but the tree inferred from the data set with third codon positions included was better resolved with higher node supports ( Figure 4; Figure S1 View Figure 1 ). This latter analysis suggests a possible 20 putative species ( Figure 4), whereas only16 were inferred
by the ABGD analyses ( Figure S2 View Figure 2 ). Nevertheless, concerning the tropical IWP species, the focus of the current paper, both Bayesian COI analysis and ABGD were congruent in rendering 10 lineages.
The tropical Indian Ocean species H. natalensis represented by specimens from South Africa, United Arab Emirates and India received maximum support maximum support measured as posterior probabilities (PP) ( Figure 4, PP = 1) and was rendered sister to the West Pacific H. japonica Pilsbry, 1895b (also known to be invasive in the East Pacific coast of the US and Mediterranean Sea; Álvarez et al. 1993; Gosliner and Behrens 2006; Hanson et al. 2013) ( Figure 4, PP = 1).
Specimens of the colourful IWP species H. ovalis and H. cymbalum clustered together ( Figure 4, PP = 1), but the results suggest three species, namely H. cymbalum proper with representatives from the Philippines, Indonesia and Lakshadweep Islands ( India) as well as two cryptic species of H. cf. ovalis , one formed by specimens from the Philippines and Andaman Islands ( India), and the other one with a single specimen from Hawaii.
The remaining four IWP species formed a clade with maximum support ( Figure 4, PP = 1). This clade includes specimens from the Indian Ocean that match the description of H. pemphis ( Philippi, 1847) , here provisionally ascribed to the latter species which has its type locality in the Red Sea, two additional species represented by western Pacific specimens resulting from GenBank sequences ( Haminoea sp. 1 and Haminoea sp. 2 ), and a fourth species here described as new ( Haminoea aptei sp. nov.) based on specimens from the Andaman and Lakshadweep Islands off the western and eastern coasts of India.
Concerning only the IWP species, intraspecific COI genetic uncorrected p -distance varied between 0.25% for H. cf. ovalis ( India, Indonesia) and H. cymbalum and 2.8% for H. cf. pemphis . The new species here described, Haminoea aptei sp. nov. (see Taxonomic section), is 9.1% distant from its closest relative in the tree, namely the Western Pacific Haminoea sp. 1 from Indonesia and East Timor, and 18.2% distant from the chromatically similar Western Indian Ocean and sympatric species H. natalensis ( Figure 4 and Table 3).
| BNHS |
Bombay Natural History Society |
| MH |
Naturhistorisches Museum, Basel |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |