
Abelisauridae Bonaparte & Novas, 1985
|
publication ID |
https://doi.org/10.11646/zootaxa.3759.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:E9F2EADE-3745-468B-9344-1ECCA4C2EA22 |
|
DOI |
https://doi.org/10.5281/zenodo.5038696 |
|
persistent identifier |
https://treatment.plazi.org/id/03C38794-FFFA-0806-53E4-FE8ABDAFF89D |
|
treatment provided by |
Felipe |
|
scientific name |
Abelisauridae Bonaparte & Novas, 1985 |
| status |
|
Abelisauridae Bonaparte & Novas, 1985
Gen. and sp. indet.
Referred material. ML 327 and ML 966 ( Figs. 3–4 View FIGURE 3 View FIGURE 4 ).
Locality and horizon. Cliffs of Lourinhã, Lourinhã, Portugal. Lourinhã Formation, Kimmeridgian-Tithonian, Upper Jurassic.
Description. ML 327 lacks the lowermost part of the crown, a small piece of the mesial carina on the lingual face and a few denticles on the distal carina. However, the crown is well preserved and most of the denticles are intact. The apical part of the distal carina of ML 966 is also missing; otherwise this tooth is relatively wellpreserved, with some part of the enamel cracked and missing.
Crown. The teeth are slightly elongated baso-apically (CHR of 1.58 in ML 327 and 1.95 in ML 966) and ziphodont in shape. Both crowns are only weakly curved distally, and the apex has been worn.
In lateral view, the distal carina is slightly concave, almost straight. The axis passing through the basal part of distal carina is perpendicular to the transversal plane of the crown. The mesial margin of the crown is much more recurved than the distal margin and the curvature is more important apically than basally. The apex is not acute and pointed but slightly rounded. In ML 327, it shows a small spalling surface on the labial face and a large wear facet ( Fig. 3A View FIGURE 3 ) corresponding to an elongated tongue-shaped surface bearing diagonal striations and inclined mesiobasally on the two-thirds of the lingual side. The spalling surface on the lingual side of the crown in ML 966 is rather subtriangular and only limited to the apex. Both mesial and distal carinae are serrated from the base to the tip of the crown. The lingual surface of ML 327 bears a prominent longitudinal depression on its mesial part, 4 mm from the mesial carina at the mid-crown. This narrow groove ( Fig. 3H View FIGURE 3 ) of 1.5 mm width extends from around 8.5 mm above the cervix dentis (or neck of the tooth, here referred to as 'cervix' sensu Smith & Dodson 2003) and ends at a distance of 8 mm from the apex. The longitudinal depression roughly follows the curvature of the crown, is closer to the mesial carina at its basal and apical endings, and almost contacts the large wear facet apically. No longitudinal groove is present on the labial face of the crown in ML 327 and on both labial and lingual sides of the tooth in ML 966.
In mesial view, the mesial carina of both teeth is concave and inclined baso-lingually. The carina remains medially positioned on the tip of the crown but twists lingually towards the root more basally and extends mesiolingually to the cervix ( Figs. 3B View FIGURE 3 and 4B View FIGURE 4 ). The crown apex remains straight and follows the general curvature of the crown. The lingual surface is slightly baso-apically sigmoid with the basal part of the crown concave and the apical one convex. On the other hand, the entire labial surface of the crown is strongly convex baso-apically. There is a flattened surface at the base of the mesial margin which is delimited lingually by the mesial carina in ML 327. This flattened surface, which appears above the cervix, extends on the first third of the crown. In ML 966 however, the surface at the base of the mesial margin is strongly convex.
In distal view, the distal carina is weakly sigmoid with a large bow oriented lingually along the basal two-thirds of the crown while the apical part of the distal carina is straight. The carina is slightly lingually positioned on the distal margin of the crown but moves medially at the tip.
In apical view, the tip of both crowns is distally positioned, with no curvature on the lingual or labial sides. The labial margin is globally convex but the distal surface is rather flattened or weakly convex. On the contrary, the surface adjacent to the distal carina on the lingual margin is rather slightly concave. In ML 327, the mesial part of the labial face is strongly convex whereas the mesial part of the lingual surface has a double curvature due to the presence of the longitudinal depression. In both teeth, the distal carina is angular whereas the mesial carina forms a low but pointed ridge which strongly displaces lingually towards the root.
In basal view, the cross-section outline of the crown base is elliptical and slightly lanceolate (i.e., mesial margin convex and distal margin pointed) in ML 327 ( Fig. 3G View FIGURE 3 ) whereas ML 966 has a well-marked lanceolate outline of the crown base ( Fig. 4G View FIGURE 4 ). In ML 327, the mesial part is roughly triangular in shape with the tip of the triangle pointed mesio-lingually whereas the mesial part of ML 966 is strongly subtriangular with the tip of the triangle medially positioned. In both crowns, the distal margin of the crown forms a semicircle. The distal margin bears the superficial ridge of the distal carina which is mesio-lingually positioned. The labio-lingual width of the base of the crown is bigger mesially (CBW of 10.69 in ML 327 and 12.94 in ML 966). With their rather flattened bases, the middles of the lingual and labial faces are almost parallel. The middle of the labial surface remains roughly flat towards the tip while the lingual surface becomes strongly convex apically. In ML 327, the dentine layer is thin ( 1 mm on the labial margin) and becomes thicker in the distal part of the crown ( 1.9 mm). Although the lingual margin has been damaged in this tooth, the pulp cavity seems to share the same lanceolate outline of the crown-base, but there is a weak labio-lingual constriction of the cavity 8 mm below the extremity of the distal carina.
Denticles. The mesial carina of ML 327 bears 11 denticles per 5 mm at the tip, 13 at the mid-crown and 20 near the cervix. In ML 966, the mesial carina shows 15 denticles at mid-crown, and 19 denticles at the base, the mesioapical denticles having been worn off. ( Table 3 View TABLE 3 ). In both crowns, the denticles decrease in size towards the root at two-thirds of the crown and the most basal denticles are minute. In lateral view, the mesial denticles are longer baso-apically than mesio-distally, which give them a subrectangular (or 'cartouche-like' sensu Harris 1998) outline. Since the denticles are inclined towards the tip of the crown and the main axis of the denticle is not perpendicular to the mesial margin of the crown, the shape of the denticle is rather parallelogram-shaped. The external margin of the mesial denticles is rounded and sometimes asymmetrically convex, with the concavity positioned slightly apically. In both teeth, the lingual and labial surfaces of the denticles are convex and the interdenticular space is shallow. In mesial view, the denticles are not labio-lingually large, they are roughly chisellike in shape but their external margin is rounded, and the main body of the denticles is almost cylindrical. There is no interdenticular sulcus between the mesial denticles in both teeth.
The distal carina of ML 327 has 12 denticles per 5 mm at the apex, 13 at the mid-crown, and around 15 at the crown base (but not near the cervix, this part being missing) so that they are similar in size to mesial denticles (DSDI of 1.14). In ML 966, 14, 12 and 14 denticles per 5 mm can be observed at the apex, mid-crown and base of the crown, respectively, and this tooth also share a DSDI close to one ( Table 3 View TABLE 3 ). Unlike the mesial denticles, the distal denticles of both crowns are longer mesio-distally than baso-apically, except in the apical denticles which are squared-like in shape, and the main axis of the denticles is perpendicular to the distal margin. In lateral view, some distal denticles of ML 327 show an external margin pointing slightly towards the tip of the crown ( Fig. 3E View FIGURE 3 ), so that the apical margin of the denticles is weakly concave whereas the basodistal margin is strongly convex. In all other distal denticles of ML 327 and all distal denticles of ML 966, the external margin is asymmetrically convex, with the denticle apex slightly apically positioned ( Fig. 4I View FIGURE 4 ). In both teeth, the labial and lingual surfaces of the denticle body are convex. The distal denticles also have a deeper interdenticular space than the mesial ones and their external margin is more acute, giving them a real chisel-like shape in distal view. In ML 327, the enamel layer is thicker than in the mesial denticles and, in both teeth, most of denticles show an elongated interdenticular sulcus diagonally oriented basally away from the denticles ( Figs. 3E View FIGURE 3 and 4E View FIGURE 4 ). These shallow grooves are parallel to each other and extend from the base of the interdenticular space and run on both labial and lingual faces of the crown. They are shorter in the apical denticles, and completely absent in the most apical one, both on the labial and lingual surface. Their inclination also tends to be reduced towards the root with interdenticular sulci being almost perpendicular to the distal margin in the basal denticles.
Surface. The enamel surface of both crowns is very well preserved and shows perfectly a granular and irregular texture on both lingual and labial faces. Besides the large longitudinal depression present on the lingual face, transversal and shallow undulations are present on both lingual and distal surfaces in ML 327 ( Fig. 3I View FIGURE 3 ). On the labial face of this tooth, they form large parabolic furrows curving apically near the distal carina, disappearing on the mesial part of the labial face due to the strong curvature of the crown. On the lingual face of this crown, they are visible distally, near the distal carina, and also in the middle of the crown, in the mid-crown surface. The undulations are absent on both convex surface adjacent to the mesial carina and the longitudinal depression. Unlike the labial wrinkling, these undulations do not bent towards the tip of the crown near the carina. In ML 966, the transversal undulations are also clearly visible on both sides of the crown ( Fig. 4A View FIGURE 4 ). They are particularly pronounced close to the distal carina on the labial margin where they also curve apically adjacent to the distal carina ( Figs. 4E, H View FIGURE 4 ). As in ML 327, the transversal undulations are large, parabolic and shallow on the lingual side of the crown, and they do not curve toward the apex close to the carinae. In both teeth, these undulations are parallel and irregularly spaced and there are approximately 3 to 4 wrinkles per 5 mm on both faces of those crowns.
Discussion. Since the root is absent, ML 327 and ML 966 are most likely shed teeth. The labio-lingual compression of these moderately large teeth ( CH > 30 mm), associated with serrated mesial and distal carinae and curvature of the tip distally, is a plesiomorphic condition seen in theropod dinosaurs. Among known large terrestrial Jurassic groups of vertebrates, this combination of characters is only seen in theropods.
Although ML 966 is slightly bigger than ML 327 ( Table 3 View TABLE 3 ), both teeth can confidently be associated to the same taxon as they share the same outline, CBR, DSDI, and the following features: presence of well-developed interdenticular sulci pointing basally, transversal undulations on both labial and lingual faces, a mesial carina offset, strongly twisted lingually towards the root and reaching the cervix, a distal carina slightly sigmoid and lingually positioned, a lingual face baso-apically concave and a labial surface baso-apically sigmoid, and a lanceolate outline of the base-crown in cross-section. Nevertheless, some denticles of ML 327 differ from ML 966 as their external margins are pointing apically and are not asymmetrically convex on their entire distal margins. However, denticle recurvature can vary in tooth row ( Fanti & Therrien 2007; see below). The interdenticular space of the distal denticles is wider in ML 966, and the crown is also slightly more elongated than ML 327 (CHR of 1.95 and 1.58) but elongation of the crown also varies greatly along the tooth row in theropods (e.g., Ceratosaurus , Allosaurus , Proceratosaurus , Tyrannosaurus ).
One of the most striking features in these two isolated teeth is the presence of tenuous to well-marked transversal undulations ('enamel wrinckles' sensu Brusatte et al. 2007) on the crown. Thought to be a possible tetanuran synapomorphy (Brusatte et al. 2007), transversal undulations are present on the crown of many theropods, from basal to derived forms, as well as metriorhynchid crocodylomorphs ( Andrade et al. 2010) and rauisuchian crurotarsans ( Brusatte et al. 2009b), and this feature cannot therefore be considered as a reliable tool alone for identifying teeth. In theropods, they have indeed been observed in basalmost theropods such as Sanjuansaurus gordilloi (PVSJ 605) and Eodromaeus murphi (PVSJ 561), ceratosaurs such as Ceratosaurus nasicornis (USNM VP 4735), Berberosaurus liassicus (MNHN Pt369), Genyodectes serus (MLP 26–39), Abelisaurus comahuensis (MPCA 1, 229, 687), Aucasaurus garridoi (MCF-PVPH 236) and Majungasaurus crenatissimus (FMNH PR 2278) , all non-Maniraptoriformes tetanurans (see Brusatte et al. 2007), and some deinonychosaurs like Troodon formosus (DMNH 22337) and Dromaeosaurus albertensis (AMNH 5356).
ML 966 also displays pronounced undulations adjacent to the distal carina. Short and marginal undulations close to carinae are a well-known feature of carcharodontosaurids teeth ( Sereno et al. 1996; Coria & Currie 2006) as they appear on the teeth of Carcharodontosaurus saharicus (SGM Din-1; UC PV6), Mapusaurus roseae (MCF- PVPH 108) and Giganotosaurus carolinii (MUCPv-CH-1). However, marginal undulations have also been reported among non-carcharodontosaurid theropods such as the abelisaurid Skorpiovenator bustingorryi ( Canale et al. 2009) . They actually seem to be present in a large range of non-coelurosaur averostrans as they have also been noticed in other ceratosaurs such as Ceratosaurus nasicornis (USNM 4735), Abelisaurus comahuensis (MPCA 5) and Majungasaurus crenatissimus (FMNH 2100) , megalosaurids like Afrovenator abakensis (UC UBA1), Megalosaurus bucklandii (NHM R.234; OUMNH J.23014) and Torvosaurus tanneri (ML 1100) , spinosaurids such as Baryonyx walkeri (NHM R.9951), Suchomimus tenerensis (MNN G35-9), and Irritator challengeri (SMNS 58022), and other allosauroids like Allosaurus fragilis (USNM 8335), Neovenator salerii (MIWG 6348) and Acrocanthosaurus atokensis (NCSM 14345).
Both teeth also possess a slightly curved distal profile of the crown, with the apex of the teeth located just apical to the most distal point of the crown at the cervix. This feature was considered to be a potential synapomorphy for Abelisauridae by Smith (2007) as a straight or slightly curved distal profile of the crown exists in Majungasaurus crenatissimus , Indosuchus raptorius , Rugops primus , Kryptops palaios , Aucasaurus garridoi (Smith & Vechia 2006; Smith & Lamanna 2006; Smith 2007; Candeiro 2007; pers. obs.) and many indeterminate abelisaurids (e.g., UCPC 10; MNHN MRS 1619, MRS 1620). Although the distal profile of the crown displays a strong curvature in most other theropods ( Ezcurra 2009; pers. obs.), a weak curvature of the distal profile can also occur in some teeth of basalmost theropods (PVSJ 512), ceratosaurids (USNM 4735; MLP 26-39), noasaurids (PVL 4061), allosauroids (SGM Din1; MCF-PVH 108.43), tyrannosauroids (MIWG 1997.550; USNM 12814; FMNH PR 2081) and some coelurosaurs ( Currie et al. 1990: fig. 8.5A; Sankey et al. 2002: fig. 4.10); therefore, the systematic utility of this feature requires association with other characters.
Nevertheless, the presence of strongly developed and elongated interdenticular sulci between distal denticles seems to be a condition genuinely shared by non-maniraptoriform averostrans. This feature has been observed in the abelisaurids Kryptops palaios (MNN GAD1−1) and Majungasaurus crenatissimus (FMNH PR 2100, 2278), the megalosauroid Piatnitzkysaurus floresi (PVL 4073), the megalosaurids Megalosaurus bucklandi (OUMNH J13506 View Materials ) and Torvosaurus tanneri (ML 1100) , the carcharodontosaurids Giganotosaurus carolinii (MUCPv-CH-1) and Mapusaurus roseae (MCF-PVPH-108), and the tyrannosaurid Tyrannosaurus rex (FMNH PR2081) . However, an irregular texture of the enamel (i.e., no specific orientation of the enamel wrinkling texture) seems to be present in most non-tetanurans theropods such as Coelophysoidea and Abelisauroidea, some tyrannosaurids and many primitive coelurosaurs, Compsognathidae and Deinonychosauria (pers. obs.). On the other hand, a braided/veined oriented texture of the enamel has been observed in Ceratosauridae, Megalosauroidea, Allosauroidea and Tyrannosauroidea and it is therefore unlikely that ML 327 and ML 966 belong to one of those clades.
A peculiar anatomical feature of ML 327 is also the presence of distal denticles with an apex pointing towards the tip, a feature present in the teeth of some abelisauroids such as Masiakasaurus knopfleri (FMNH PR 2221, 2296), Kryptops palaios (MNN GAD1−1), Rugops primus (MNN IGU1), Majungasaurus crenatissimus (FMNH PR 2008, 2100, 2278) and other abelisaurid taxa (e.g., MUCPv 482; MUCPv 641). Among large theropods like ceratosaurids, megalosauroids, allosauroids and tyrannosauroids, the denticles are symmetrically rounded or slightly asymmetrically convex in lateral view but never hooked apically ( contra Bakker & Bir 2004 for ceratosaurids and allosaurids, and Smith 2007 for tyrannosaurids; Currie et al. 1990; Abler 1992; pers. obs.). Slightly to strongly hooked distal denticles can also be observed in the basal saurischian Eoraptor lunensis (e.g., third right premaxillary tooth; PVSJ 512) and many Troodontidae (e.g., Currie 1987; Currie et al. 1990; Holtz et al. 1998; Longrich 2008; pers. obs.) and Dromaeosauridae (e.g., Currie et al. 1990; Currie & Varricchio 2004; Baszio 1997; Longrich 2008; pers. obs.). Deinonychosaurs, however, possess either very large and well-separated serrations, as in troodontids and Saurornitholestes , or a number of denticles per five millimeters higher than 14 on the distal carina ( Smith et al. 2005). Likewise, both dromaeosaurids and Masiakasaurus tend to have distal denticles larger to mesial serrations ( Currie et al. 1990; Currie & Varricchio 2004; Norell et al. 2006; Longrich 2008; pers. obs.). To our knowledge, neither noasaurids nor deinonychosaurs display a combination of pronounced and elongated interdenticular sulci and short marginal undulations on the crown.
Interestingly, ML 966 lacks hooked denticles on the distal carina as all denticles are either symmetrically or asymmetrically convex. This would therefore suggest that apically recurved denticles might not be present in all teeth along the tooth row. Denticle recurvature seems indeed to vary in the dentition of Majungasaurus crenatissimus as strongly recurved denticles are present in lateral and mesial dentary teeth and slightly recurved to symmetrically rounded denticles exist in some lateral and premaxilla teeth ( Fanti & Therrien 2007; pers. obs.).
The presence of an elongated and deep groove adjacent to the mesial carina on the lingual side of the crown in ML 327 is another peculiar feature that, to our knowledge, has not been observed in any teeth belonging to a large theropod (crown with CH > 30 mm), and might therefore represent an autapomorphy. A concave surface adjacent to the mesial carina can be observed in the mesialmost teeth of many abelisaurids such as Rugops primus (MNN IGU1), Indosuchus raptorius (AMNH 1753) and Majungasaurus crenatissimus (FMNH PR 2100) , but also in Allosaurus fragilis (AMNH 851), some tyrannosauroids such as Proceratosaurus bradleyi (NHM R 4860) and Eotyrannus lengi (MIWG 1997.550), and some dromaeosaurids like Dromaeosaurus albertensis (AMNH 5356). However, the surface adjacent to the mesial carina in ML 327 is convex and the concave area formed by the longitudinal groove is narrow. Longitudinal grooves running along the crown surface can also be observed in several theropod taxa such as Scipionyx samniticus ( Dal Sasso & Maganuco 2011) , Buitreraptor gonzalezorum and Austroraptor cabazai ( Gianechini et al. 2011b) , and there are two grooves separated by a large medial ridge ( Gianechini et al. 2011a; Gianechini et al. 2011b; pers. obs.). Likewise, the mesial groove present in ML 327 cannot be confused with the large medial concavity (‘supradental groove' of Gong et al. 2010) present on the crown of many theropods like Orkoraptor burkei ( Novas et al. 2008) and Sinornithosaurus ( Gong et al. 2010) , or the numerous flutes visible on the teeth of Coelophysis bauri ( Buckley 2009) , Masiakasaurus knopfleri ( Carrano et al. 2002) , Ceratosaurus nasicornis ( Madsen & Welles 2000) , spinosaurids (e.g., Charig & Milner 1997; Sereno et al. 1998; Sues et al. 2002), Paronychodon lacustris (e.g., Cope 1876; Sankey et al. 2002; Baszio 1997; Sankey 2008) or Velociraptor mongoliensis (AMNH 6515).
On the basis of the combination of several important features in ML 966 and ML 327, a large crown ( CH > 30 mm), an almost straight distal profile of the tooth, transversal and short marginal undulations on the crown, denticles with strongly developed interdenticular sulci, a DSDI close to one, an irregular enamel texture and the presence of apically pointed denticles on the distal carina in ML 327, these two teeth are assigned to a member of the Abelisauridae . Within this clade, ML327 and ML 966 only differ from other abelisaurids by having a strongly twisted mesial carina. However, this feature is also present in some basal abelisaurids such as Abelisaurus (MPCA 685). Also, ML 327 has a labially displaced distal carina which contrasts with the centrally positioned carina on the distal margin of the crown of abelisaurids (pers. obs.).
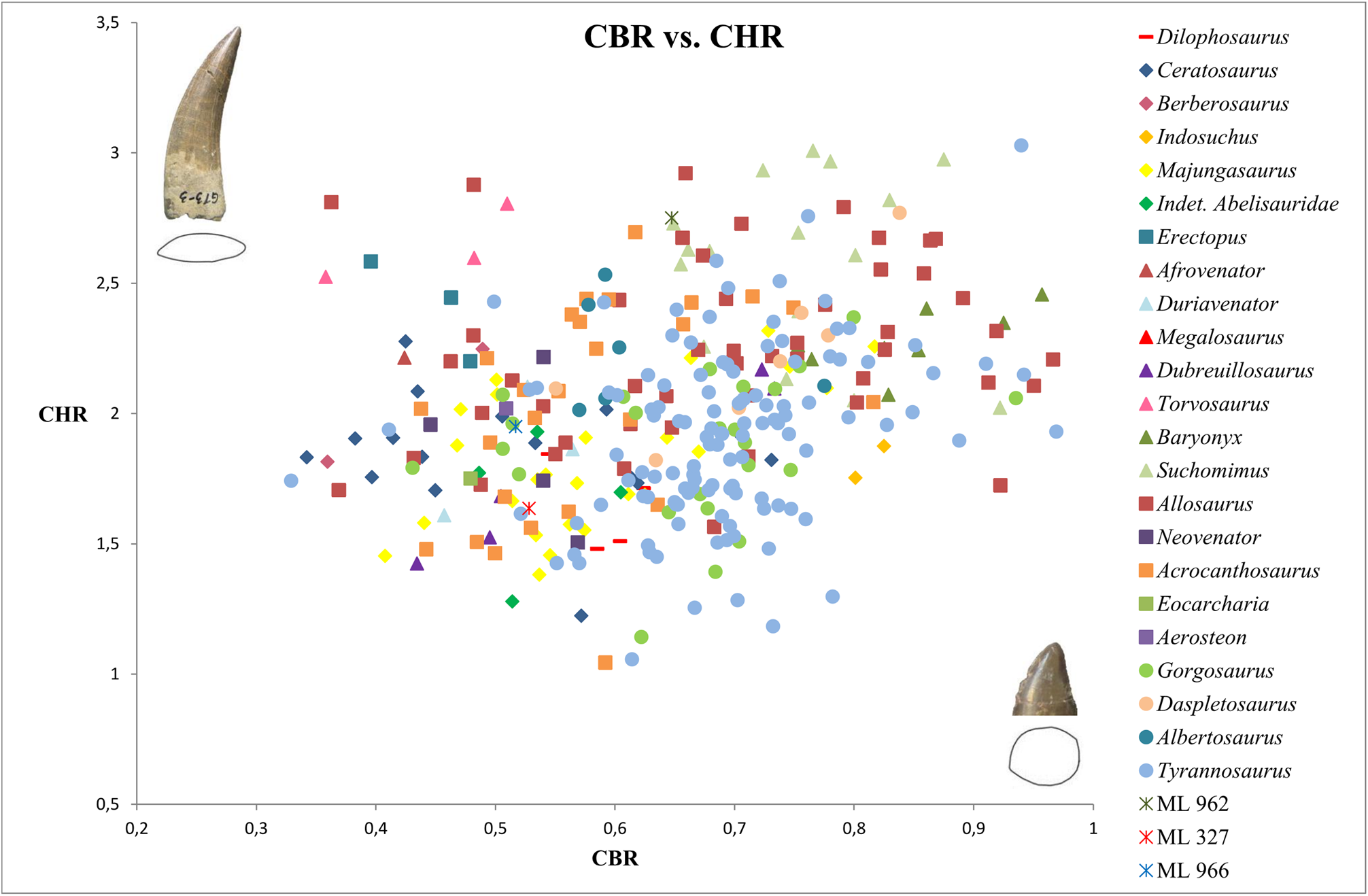
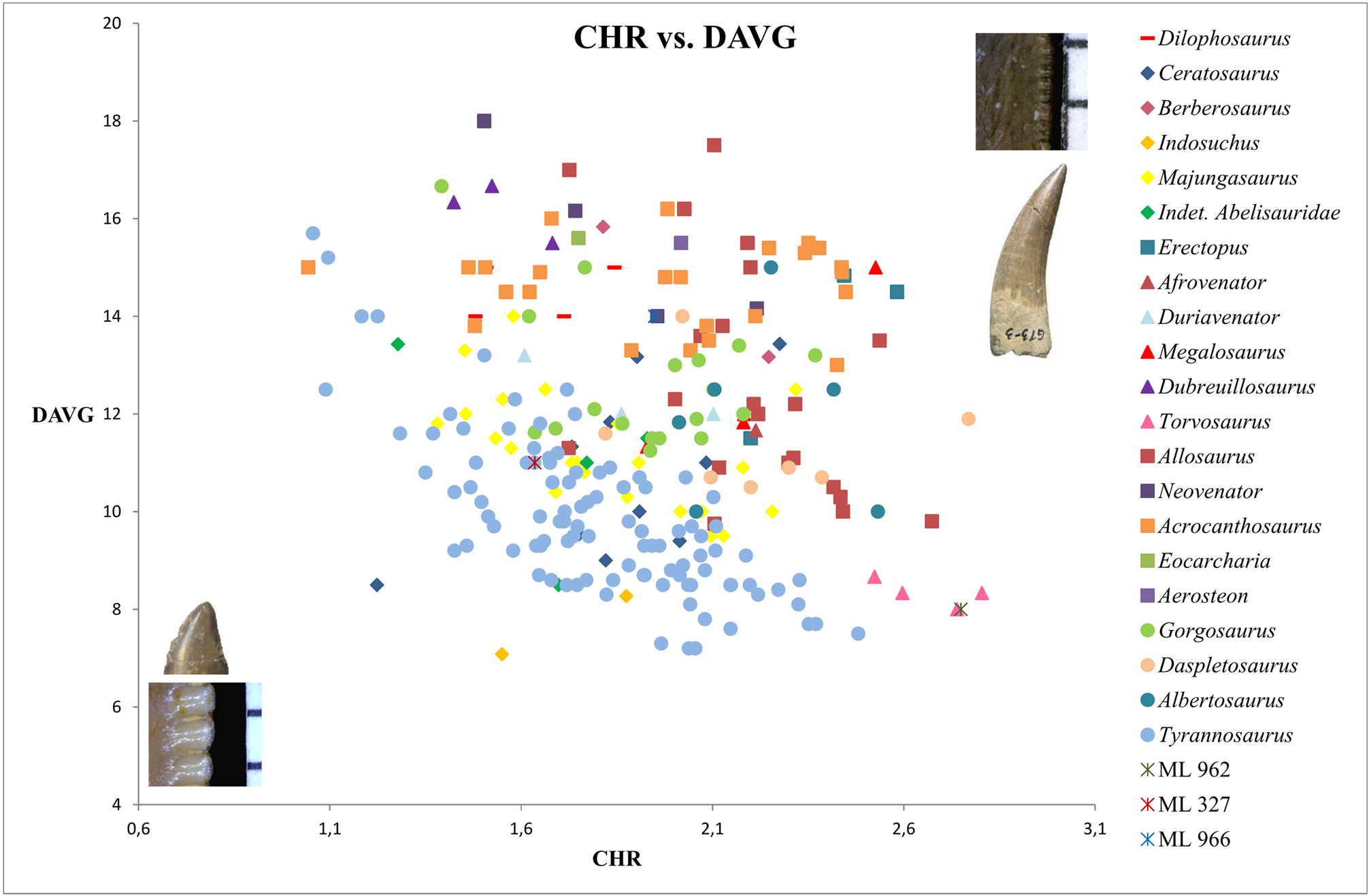
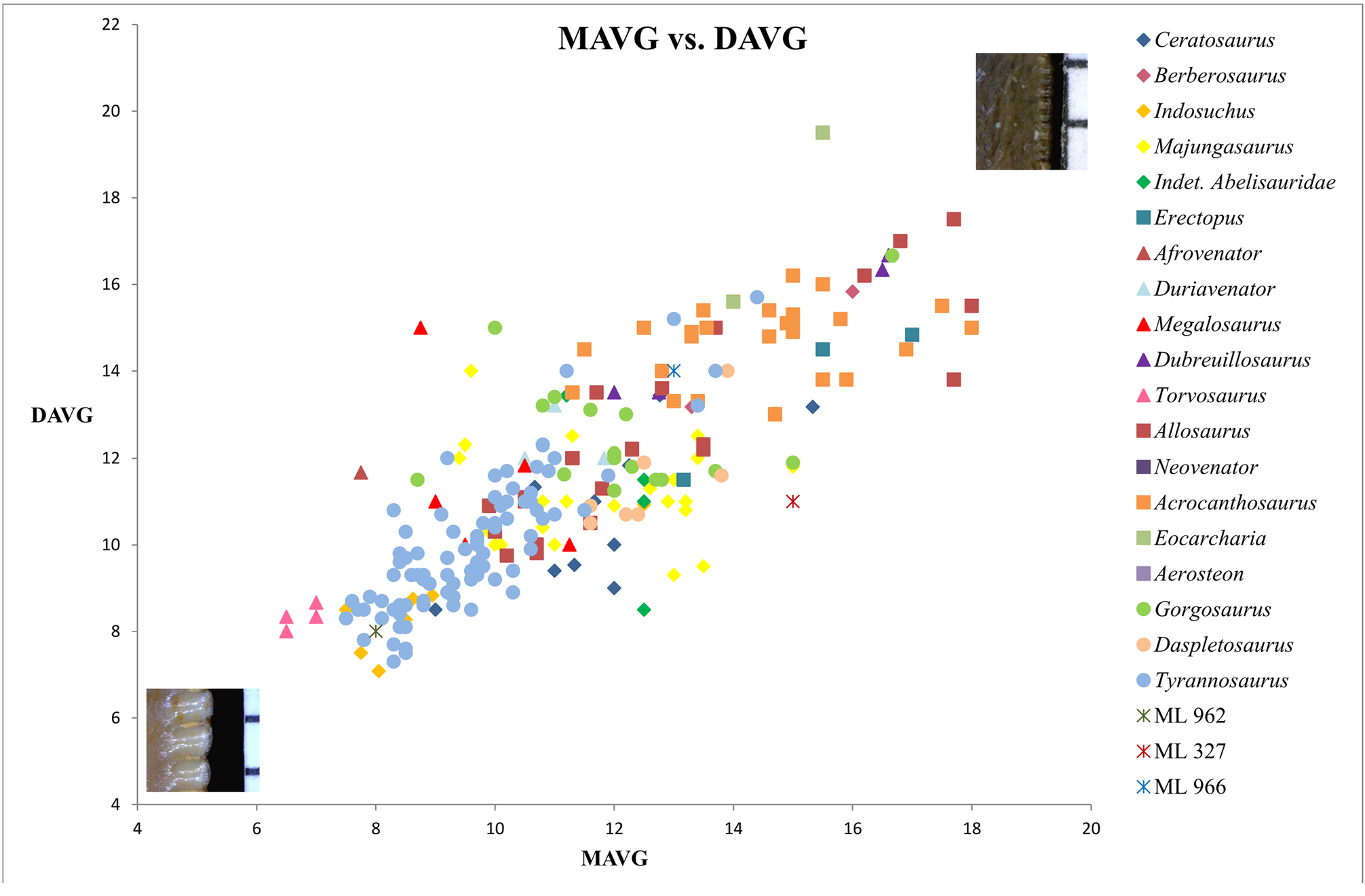
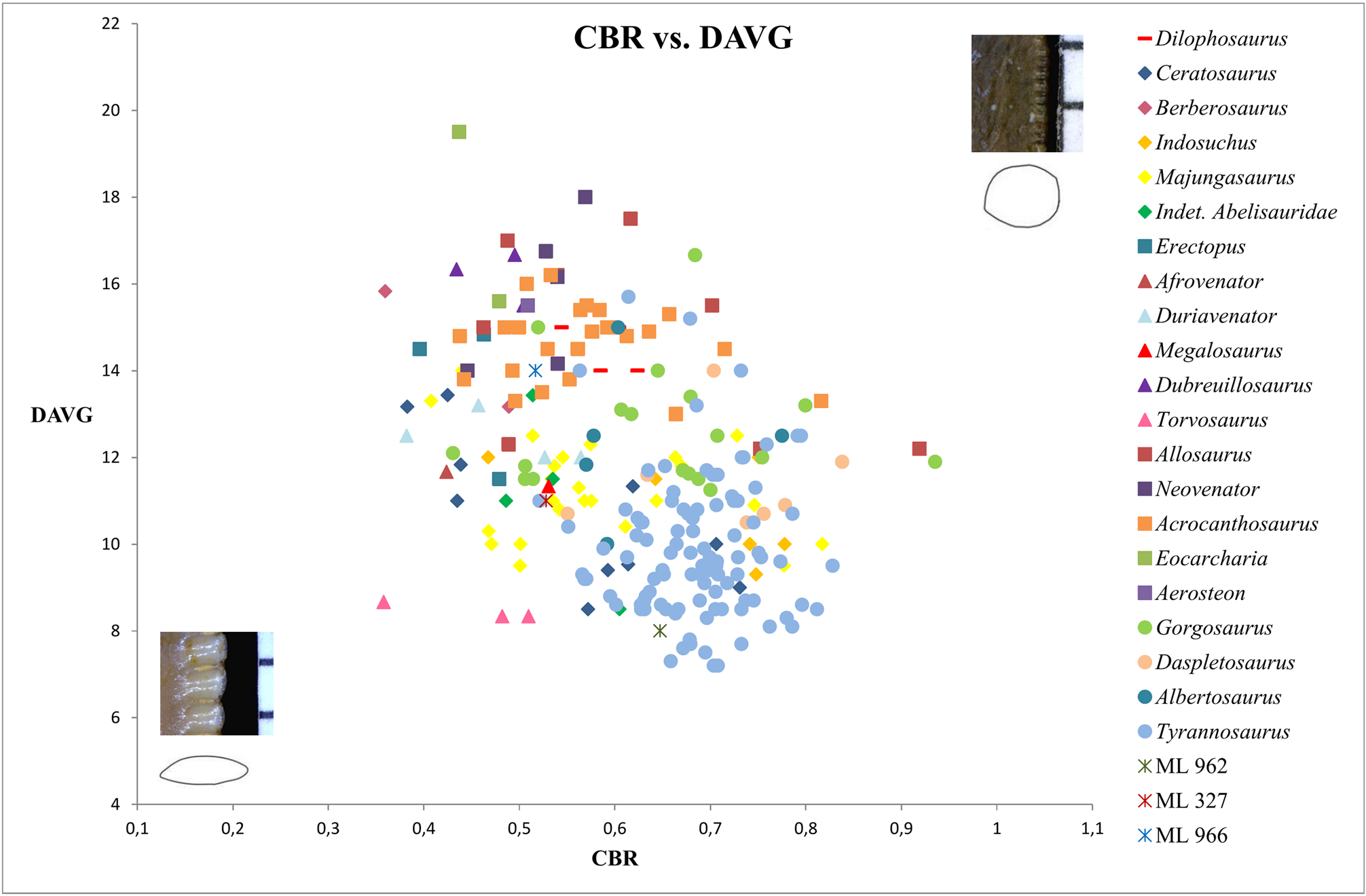
Bivariate plots of CBR and CHR reveal that ML 966 and ML 327 mainly occupy the same area of values as Abelisauridae ( Majungasaurus + indeterminate abelisaurids), Ceratosaurus , Allosaurus , Acrocanthosaurus and Gorgosaurus teeth ( Fig. 5 View FIGURE 5 ). However, bivariate plots with MAVG or DAVG clearly show that the two teeth possess smaller mesial and distal denticles than any abelisaurids represented, with a number of denticles per five mm situated among the values of Allosaurus , Acrocanthosaurus and Berberosaurus ( Figs 6–8 View FIGURE 6 View FIGURE 7 View FIGURE 8 ). The number of denticles per five mm of ML 966 and ML 327 are indeed situated between 13 to 16, a higher number than in Majungasaurus , Indosuchus , Rugops and UCPC 10 ( Smith 2007; Sereno & Brusatte 2008; pers. obs.) but comparable to that of the most basal abelisaurid Kryptops ( Sereno & Brusatte 2008) and Abelisaurus (pers. obs.).
Due to the relatively important labiolingual compression of the base crown (CBR close to 0.5), ML 966 and ML 327 are most likely lateral teeth and have therefore been coded as such in our datasets. When the two isolated teeth are included in the dentition-based data matrix, the resulting consensus tree of the cladistic analysis retrieved both teeth together in a well-supported clade (Bremer support of 4) nested among abelisaurid theropods ( Fig. 1 View FIGURE 1 ). Both isolated teeth form the sister taxon of a clade encompassing the abelisaurids Rugops , Kryptops and Majungasaurus , and the monophyletic group formed by ML 966, ML 327 and these three abelisaurids is supported by two ambiguous synapomorphies: the long and well-developed interdenticular sulci of basal and mid-crown denticles on the distal carina (char. 105 and 106). When incorporated into the supermatrix, the cladistic analyses resulted in a poorly resolved consensus tree in which ML 327 and ML 966 were found as sister-taxa among the clade of Abelisauridae (Appendix, Fig. A10 View FIGURE 10 ). The deletion of the wildcard taxa Erectopus and Piatnitzkysaurus resulted in a better resolved consensus tree in which ML 327 and ML 966 are still nested in the same clade within Abelisauridae ( Fig. 2 View FIGURE 2 ).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |