
Corynactis carnea Studer, 1879
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4688.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:40A12FD4-A0FA-4AE6-AAC6-6E92280A8608 |
|
persistent identifier |
https://treatment.plazi.org/id/03C38D60-FFBF-4C55-ADA4-FED658E4B727 |
|
treatment provided by |
Plazi |
|
scientific name |
Corynactis carnea Studer, 1879 |
| status |
|
Corynactis carnea Studer, 1879 View in CoL
Corynactis carnea Studer 1879 View in CoL ; Andres 1883; Andres 1884; Kwietniewski 1895; Duerden 1896; Haddon 1898; Duerden 1900; McMurrich 1904; Stephenson 1922; Carlgren 1927; Carlgren 1938; Carlgren 1941; Carlgren 1949; Carter Verdeilhan 1965; Riemann-Zürneck 1979; den Hartog 1980; Riemann-Zürneck 1986; den Hartog et al. 1993; Acuña & Zamponi 1995; Genzano et al. 1996; Häussermann & Försterra 2005; Rodríguez et al. 2007; Acuña & Garese 2009; Häussermann 2009; Garese et al. 2016; Fautin 2016.
Anemonia variabilis McMurrich 1893 ; Carlgren 1896; Pax 1907.
Anemonia carnea: McMurrich 1893 .
Sphincteractis sanmatiensis Zamponi 1976 ; Riemann-Zürneck 1986; Zamponi & Acuña 1992; Acuña & Zamponi 1995; Excoffon & Acuña 1995; Genzano et al. 1996; Zamponi et al. 1998 a; Zamponi et al. 1998 b; Häussermann & Försterra 2005; Fautin et al. 2007; Rodríguez et al. 2007; Lauretta et al. 2009; Häussermann 2010; Fautin 2016.
Corynactis sanmatiensis: den Hartog et al. 1993 View in CoL ; Fautin 2016.
Studied material. MLP 8504 View Materials : Sphincteractis sanmatiensis , type (see discussion regarding this subject). Seven specimens in two jars (4+3) .
MACN-IN 42230 : several specimens. Punta Este , Chubut, Argentina 42º47.141’S 64º57.282’W, intertidal; coll. Daniel Lauretta, October 2007 GoogleMaps .
MACN-IN 42227 : nine specimens . SAO, Argentinean exclusive economic zone, O/V “Puerto Deseado” Mejillón II expedition 38º14.278’S 57º09.313’W (St. 3), 51.4 m depth GoogleMaps ; coll. Daniel Lauretta, September 2009.
MACN-IN 42228 : eight specimens . SAO, Argentinean exclusive economic zone, O/V “Puerto Deseado” Mejillón II expedition 39º01.384’S 58º02.146’W (St. 11), 140 m depth GoogleMaps ; coll. Daniel Lauretta, September 2009.
MACN-IN 42229 : about 20 specimens . SAO, Argentinean exclusive economic zone, O/V “Puerto Deseado” Mejillón II expedition 36º39.868’S 55º09.779’W (St. 22), 50 m depth GoogleMaps ; coll. Daniel Lauretta, September 2009.
Description. External anatomy. Oral disc circular and flat, wider than column (preserved), in live specimens much wider than column and pedal disc ( Fig. 2 View FIGURE 2 ). Transparent or same color as column ( Fig. 2a,b,c View FIGURE 2 ). Usually a more intense color ring on the margin (green, orange). Mesenterial insertion clearly visible. Mouth central, circular or oval, strongly elevated. Biggest specimen with 94 tentacles in up to four cycles (but many irregularities occur), in rows, to 4 (5?) in one endocele. Exocelic marginal tentacles bigger than endocelics. Stalk rugose due to the presence of cnidocysts batteries. External tentacles much longer than internal (to 2.1 mm and 0.5 respectively, preserved).All with well-developed acrospheres. Oral disc can cover the tentacles completely. Column smooth; fluorescent green, light violet, orange-brown or transparent ( Fig. 2 View FIGURE 2 ). Taller than wider (to 8.85 mm and 5 mm respectively, preserved). Mesenterial insertions visible, sometimes internal features also visible. Pedal disc usually round or elliptical, firmly attached to substratum and wider than column.
Internal anatomy. Longitudinal muscles of tentacles and oral disc ectodermal ( Fig. 3a,b,d View FIGURE 3 ). Pharynx about 1/5 (rarely 1/2) of column length (preserved). Mesogloea thicker than other layers (to 110 µm at pharynx level), thicker in distal column than in proximal column, pedal and oral disc. Up to 64 mesenteries in 3 cycles (8+8+16=32 pairs), last cycle imperfect. Many irregularities found. Same number of mesenteries in proximal and distal part of column. All oldest mesenteries and some younger fertile. One siphonoglyph attached to a pair of directives. Retractor muscles diffuse, weak ( Fig. 3f View FIGURE 3 ). No basilar nor parietobasilar muscles ( Fig. 3e View FIGURE 3 ). Sphincter endodermal, diffuse, weak but clearly visible ( Fig. 3a,c View FIGURE 3 ).
Cnidae. The cnidom of C. carnea includes spirocysts (in acrospheres, tentacles, mesenterial filaments and pharynx), holotrichs (in acrospheres, column, mesenterial filaments and pharynx), microbasic b -mastigophores (in acrospheres, column, mesenterial filaments and pharynx) and microbasic p -mastigophores (in all tissues), although some cnida types are very scarce and may be contamination (see Fig. 4 View FIGURE 4 and Table 1 View TABLE 1 ).
Categories Range of length ẍ ± SD N S and width of capsules (µm) (length x mean) ACROSPHERES Spirocysts (a) (21.9–57.6) x (2.2–4.1) 37.5 ± 9.1 x 3.3 ± 0.5 36 6/6 Holotrichs (b) (65.4–87.1) x (11.8–27.1) 77.8 ± 5.3 x 16.0 ± 2.5 36 5/6 M b-mastigophores (c) (3.1–51.0) x (3.8–7.3) 38.2 ± 5.3 x 4.7 ± 1.0 20 6/6 M p-mastigophores 1 (d) (28.6–36.1) x (4.1–7.1) 31.9 ± 2.8 x 5.8 ± 1.1 12 4/6 M p-mastigophores 2 (e) (40.7–75.0) x (4.3–6.8) 57.2 ± 9.6 x 5.4 ± 0.6 19 5/6 TENTACLES Spirocysts (21.0–41.4) x (2.4–6.1) 30.7 ± 5.4 x 3.6 ± 0.7 36 4/4 M b-mastigophores* (16.5–28.9) x (4.3–4.5) - 2 1/4 M p-mastigophores* (63.3) x (6) - 1 1/4 COLUMN Holotrichs (f) (44.1–56.9) x (9.7–13.6) 50.3 ± 3.2 x 12.3 ± 1.1 28 3/6 Holotrichs (79.0–90.7) x (26.4–26.9) 85.7 ± 4.3 x 25.3 ± 2.0 5 1/6 M b-mastigophores (g) (13.8–20.6) x (2.2–4.2) 17.6 ± 2.2 x 3.3 ± 0.5 11 3/6 M p-mastigophores (h) [21.9](26.8–28.6) x (5.9–7.5) 27.5 ± 0.7 x 6.2 ± 0.7 6 4/6 M p-mastigophores (39.9–50.5) x (6.4–8.4) 45.3 ± 4.1 x 7.6 ± 0.7 7 4/6 PHARYNX Spirocysts* (30.6) x (28.9) - 1 3/6 Holotrichs (i) (34.6–50.3) x (8.6–14.0) 42.0 ± 3.5 x 11.6 ± 1.3 38 6/6 Holotrichs* (73.3–79.1) x (14.3–25.3) - 3 2/6 M b-mastigophores (j) (25.0–36.0) x (3.2–5.2) 29.6 ± 3.7 x 3.9 ± 0.5 14 4/6 M p-mastigophores (k) (21.9–37.5) x (5.6–9.2) - 3 3/6 MESENTERIAL Spirocysts* (27.2–32.1) x (2.9–3.2) - 2 4/6 Holotrichs (l) (66.9–92.3) x (19.6–30.1) 80.6 ± 6.1 x 24.2 ± 2.4 40 6/6 M b-mastigophores* (24.1–34.9) x (3.3–4.8) 27.9 ± 4.8 x 4.0 ± 0.6 4 2/6 M p-mastigophores (m) (19.6–41.1)[47.2] x (5.2–10.5) 32.8 ± 5.9 x 8.3 ± 1.3 44 6/6
FILAMENTS
Distribution and Natural History. Corynactis carnea is the most common and abundant corallimorpharian of the region. It can be found in the shallow waters of northern Patagonia, from San Matías gulf to Bahía Bustamante (Rio Negro and Chubut) and south of Buenos Aires province (in deeper waters) ( Fig. 1 View FIGURE 1 ), always associated with hard substratum, like rocks or the tubes of the polychaetes Eunice argentinensis ( Treadwell, 1929) and E. frauenfeldi Grube, 1866 (identify by S. Calla). It is also common in the Argentinean continental shelf, where it can be found up to 199 m deep ( Excoffon & Acuña 1995; Zamponi et al. 1998). It is usually found in groups of several specimens displaying the same color pattern ( Fig. 2a, d View FIGURE 2 ). Asexual reproduction present; many specimens were connected by the pedal disc. The geographic location of some specimens of Zamponi (1976) were mistakenly placed on earth: St. 17 (41º05’S; 67º07’W) and St.110 (41º13’S; 65º59’W), as noted by Riemann-Zürneck (1986).
Remarks. Corynactis carnea was first described by Studer (1879) from specimens collected off Buenos Aires ( Argentina). McMurrich (1893) described specimens from about 430 km south from Studer’s specimens under a different name, but acknowledged that both species were the same and used the name Anemonia carnea . Zamponi (1976) described Sphincteractis sanmatiensis Zamponi, 1976 from 115 specimens collected in the San Matías gulf from 16.5 to 98 m. He placed the species in Sideractidae Danielssen, 1890, a family of species with only one tentacle per endocoel and exocoel, and since the specimens present a marginal sphincter, he created a new genus Sphincteractis Zamponi, 1976 . Several works indicate with more or less confidence that S. sanmatiensis and C. carnea are the same species ( Riemann-Zürneck 1979, 1986; den Hartog et al. 1993; Rodríguez et al. 2007; Häussermann 2009). den Hartog et al. (1993) stated that C. sanmatiensis is “undoubtedly a junior synonym of C. carnea ”, but it is listed as a valid species in Fautin’s (2016) work as Corynactis sanmatiensis .
We studied the specimens of S. sanmatiensis of lot MLP 8504. The type lot (MLP 8504) is actually two different jars, with seven specimens in total. One of the jars (the one with three specimens) has a label in manuscript with the name Sphincteractis sanmatiensis and all the station numbers mentioned in the paper; there is no indication of status of the specimens (e.g. holotype, syntype). The other lot (with four specimens) has no manuscript label, but a label made with a printer that says in its left lower corner “sintipos”; this label was not made by Zamponi. One of the seven specimens is the holotype stated by Zamponi, but it is not possible to indicate which one (nor the type locality) based on the information stated in the paper or in the label. Fautin (2016) stated that the type is not a holotype as indicated in the original description, but a syntype constituted by four specimens (they are actually seven in two jars, but she probably only knew about one jar). We prefer to keep it as a holotype (as designated in the original description of the species), even when it is not possible at this moment to be sure which of the seven specimens is the holotype. The specimens are not in good conditions and it was not possible to locate the 114 (?) paratypes. We found in at least one of the specimens two tentacles in line, so we agree that Sphincteractis is a junior synonym of Corynactis . Based on the coloration pattern and anatomy described by Zamponi (1976) and cnida sizes and distribution we also agree that C. sanmatiensis is a junior synonym of C. carnea .
The internal anatomy of this species is more or less variable (e. g. number of perfect mesenteries, directives, fertile mesenteries, etc.), probably as a result of asexual reproduction. The cnidae in our specimens is in line with previous records. The cnida from one of the specimens from the lot MLP 8504 were in line with our results (although only few capsules were found). Sizes are compatible with Carlgren (1927), although no differentiation between cnida types were done (except for spirocysts). Zamponi (1976) did not report size of cnidae, but Zamponi & Acuña (1992), Acuña & Zamponi (1995) and Garese et al. (2016) did. Although there are some differences in cnida sizes, this may be due to the low number of capsules found in our specimens and variability between different specimens. Garese et al. (2016) have shown the intraspecific variation of cnida sizes in this species.
Zamponi (1976) found gametes in specimens collected between 16.5–80 meters deep (San Matías gulf); but Acuña & Zamponi (1995) did not find gametes in specimens from the intertidal zone of Puerto Madryn (Nuevo Gulf). We also failed to find gametes in our specimens from the intertidal zone, but we did find them in specimens from “Mejillón II” expedition (50–140 m). Since it was possible to find evidence of asexual reproduction in intertidal specimens, it is possible that this strategy of reproduction is preferred over sexual reproduction in those conditions, although a detail reproductive study of the species in both areas is needed to confirm this observation.
There is some hesitation about the validity of species of Corynactis from the southern hemisphere (see den Hartog et al. 1993). Since Corynactis populations of Argentina and Chile seem not to be connected, we prefer to maintain both species separated. Corynactis viridis Allman, 1846 has been reported from Cabo Frío, Brazil (den Hartog 1980; den Hartog et al. 1993). Although den Hartog et al. (1993) hesitate for geographic reasons about the affinity of these specimens with C. viridis from the northern hemisphere, they were unable to distinguish them. They indicate that if the brazilian specimens really are C. viridis then C. carnea may also be so (den Hartog et al. 1993). We agree that it is necessary to perform a study using morphological and molecular information to evaluate the validity of C. viridis , C. carnea , C. chilensis Carlgren, 1941 (separated from C. carnea by Carlgren in 1941) and C. annulata ( Verrill, 1867) .
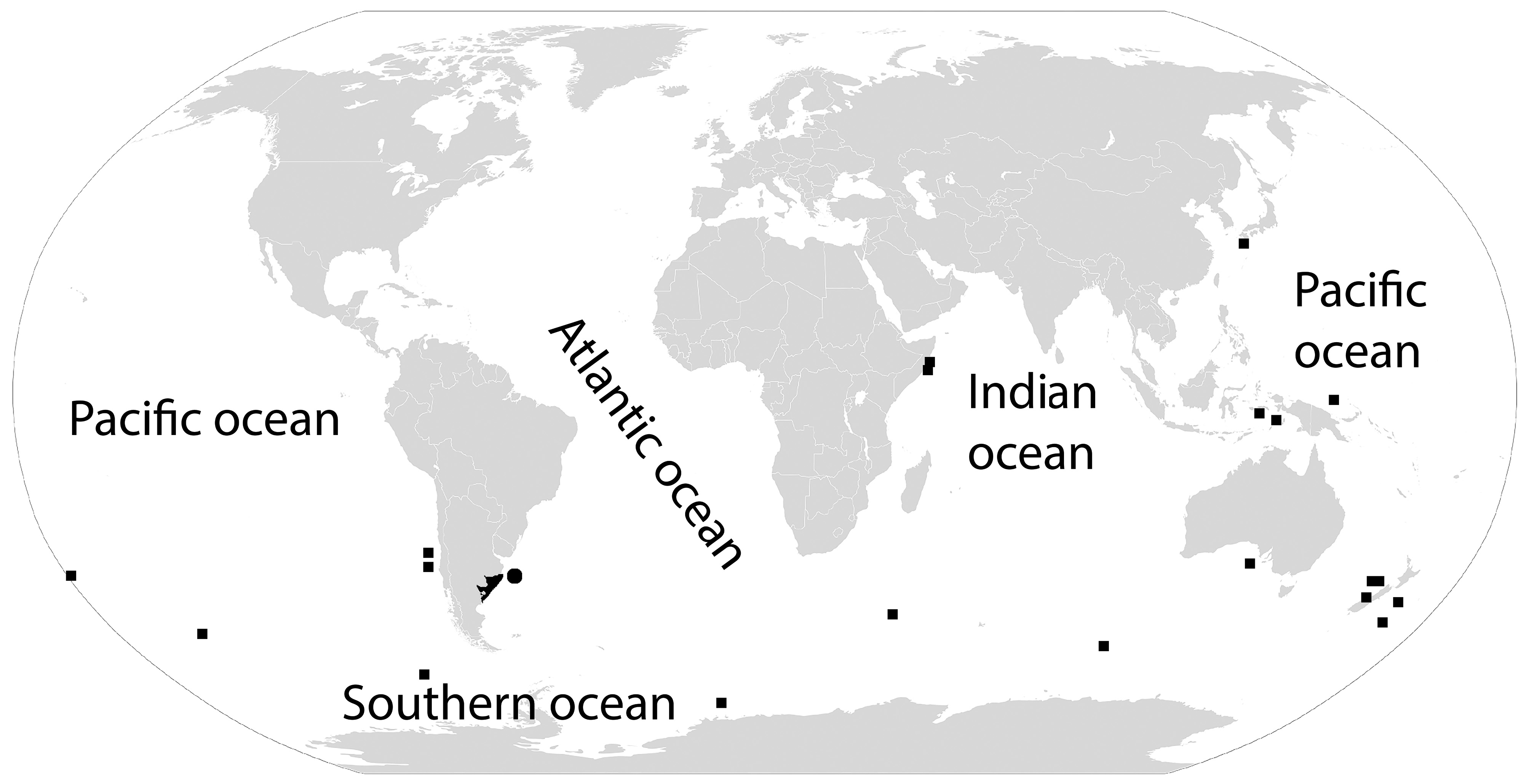

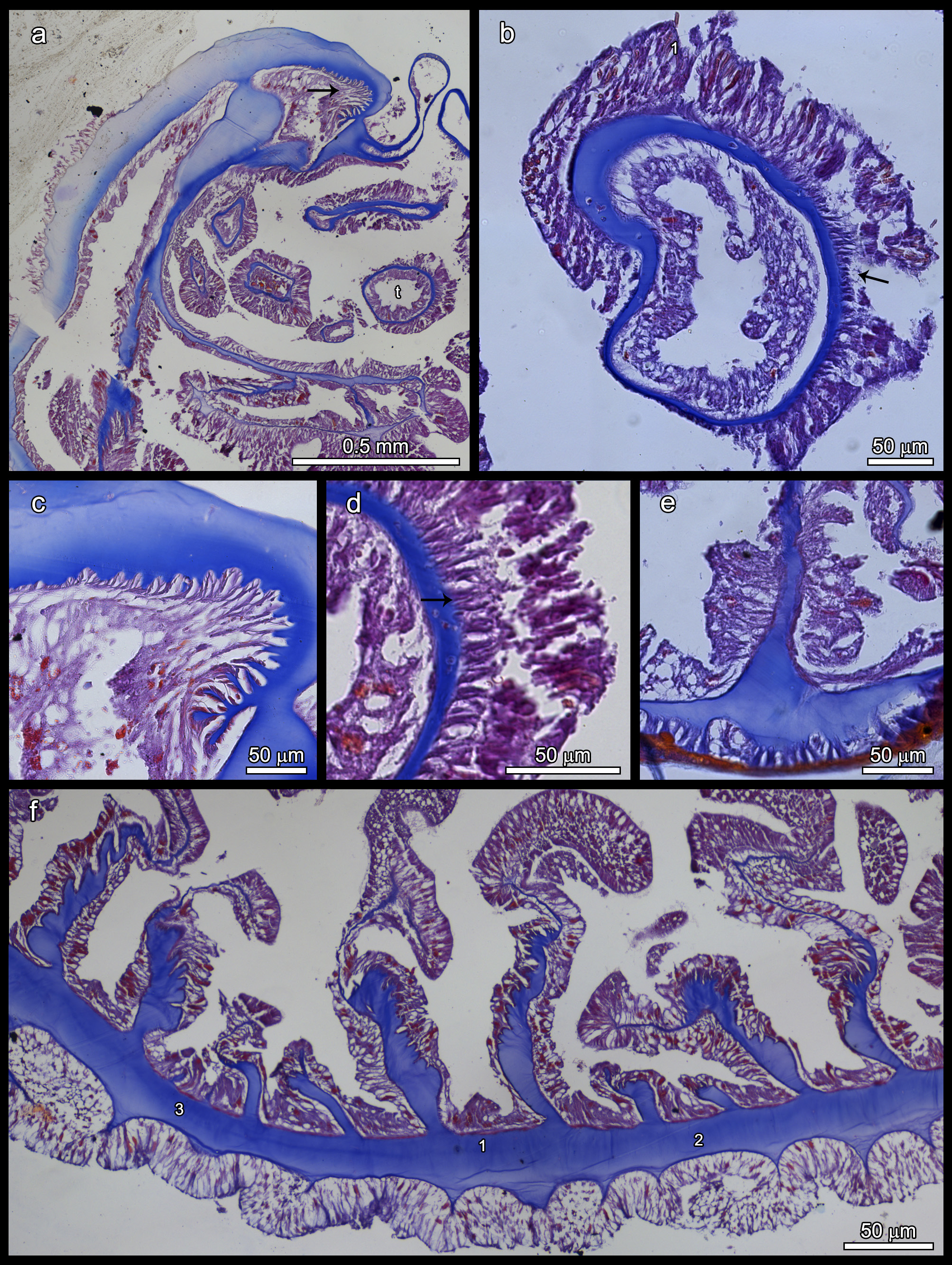

TABLE 1. Size range of the cnidae of Corynactis carnea
| Categories | Range of length | ẍ ± SD | N | S |
|---|---|---|---|---|
| and width of capsules (µm) | ||||
| (length x mean) | ||||
| ACROSPHERES | ||||
| Spirocysts (a) | (21.9–57.6) x (2.2–4.1) | 37.5 ± 9.1 x 3.3 ± 0.5 | 36 | 6/6 |
| Holotrichs (b) | (65.4–87.1) x (11.8–27.1) | 77.8 ± 5.3 x 16.0 ± 2.5 | 36 | 5/6 |
| M b-mastigophores (c) | (3.1–51.0) x (3.8–7.3) | 38.2 ± 5.3 x 4.7 ± 1.0 | 20 | 6/6 |
| M p-mastigophores 1 (d) | (28.6–36.1) x (4.1–7.1) | 31.9 ± 2.8 x 5.8 ± 1.1 | 12 | 4/6 |
| M p-mastigophores 2 (e) | (40.7–75.0) x (4.3–6.8) | 57.2 ± 9.6 x 5.4 ± 0.6 | 19 | 5/6 |
| TENTACLES | ||||
| Spirocysts | (21.0–41.4) x (2.4–6.1) | 30.7 ± 5.4 x 3.6 ± 0.7 | 36 | 4/4 |
| M b-mastigophores* | (16.5–28.9) x (4.3–4.5) | - | 2 | 1/4 |
| M p-mastigophores* | (63.3) x (6) | - | 1 | 1/4 |
| COLUMN | ||||
| Holotrichs (f) | (44.1–56.9) x (9.7–13.6) | 50.3 ± 3.2 x 12.3 ± 1.1 | 28 | 3/6 |
| Holotrichs | (79.0–90.7) x (26.4–26.9) | 85.7 ± 4.3 x 25.3 ± 2.0 | 5 | 1/6 |
| M b-mastigophores (g) | (13.8–20.6) x (2.2–4.2) | 17.6 ± 2.2 x 3.3 ± 0.5 | 11 | 3/6 |
| M p-mastigophores (h) | [21.9](26.8–28.6) x (5.9–7.5) | 27.5 ± 0.7 x 6.2 ± 0.7 | 6 | 4/6 |
| M p-mastigophores | (39.9–50.5) x (6.4–8.4) | 45.3 ± 4.1 x 7.6 ± 0.7 | 7 | 4/6 |
| PHARYNX | ||||
| Spirocysts* | (30.6) x (28.9) | - | 1 | 3/6 |
| Holotrichs (i) | (34.6–50.3) x (8.6–14.0) | 42.0 ± 3.5 x 11.6 ± 1.3 | 38 | 6/6 |
| Holotrichs* | (73.3–79.1) x (14.3–25.3) | - | 3 | 2/6 |
| M b-mastigophores (j) | (25.0–36.0) x (3.2–5.2) | 29.6 ± 3.7 x 3.9 ± 0.5 | 14 | 4/6 |
| M p-mastigophores (k) | (21.9–37.5) x (5.6–9.2) | - | 3 | 3/6 |
| MESENTERIAL | ||||
| Spirocysts* | (27.2–32.1) x (2.9–3.2) | - | 2 | 4/6 |
| Holotrichs (l) | (66.9–92.3) x (19.6–30.1) | 80.6 ± 6.1 x 24.2 ± 2.4 | 40 | 6/6 |
| M b-mastigophores* | (24.1–34.9) x (3.3–4.8) | 27.9 ± 4.8 x 4.0 ± 0.6 | 4 | 2/6 |
| M p-mastigophores (m) | (19.6–41.1)[47.2] x (5.2–10.5) | 32.8 ± 5.9 x 8.3 ± 1.3 | 44 | 6/6 |
| SAO |
Sammlung Oberli |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Corynactis carnea Studer, 1879
| Lauretta, Daniel & Martinez, Mariano I. 2019 |
Corynactis sanmatiensis:
| den Hartog 1993 |
Sphincteractis sanmatiensis
| Zamponi 1976 |
Anemonia variabilis
| McMurrich 1893 |
Anemonia carnea
| : McMurrich 1893 |
Corynactis carnea
| Studer 1879 |