
Cymbasoma annulocolle, Suárez-Morales, Eduardo & Mckinnon, David, 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4102.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:9A7BA798-AA7C-4CAA-B42C-1E260CA573E4 |
|
DOI |
https://doi.org/10.5281/zenodo.6091295 |
|
persistent identifier |
https://treatment.plazi.org/id/03C4CA6D-D507-FF80-FF12-505E96ED28AE |
|
treatment provided by |
Plazi |
|
scientific name |
Cymbasoma annulocolle |
| status |
sp. nov. |
Cymbasoma annulocolle sp. nov.
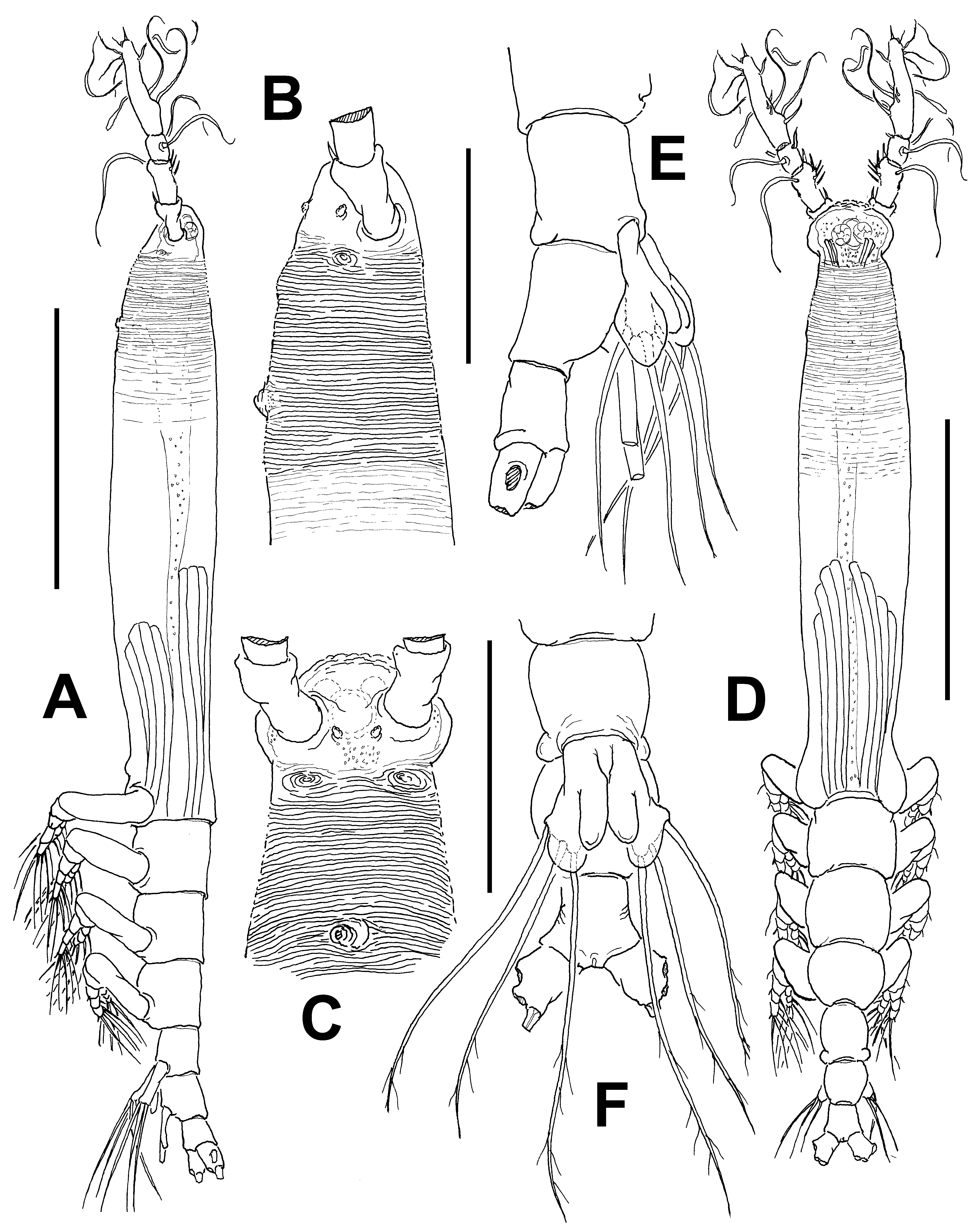
( Figs 13–19 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 View FIGURE 18 View FIGURE 19 )
Material examined. Adult female holotype, adult male allotype, two paratype females (one of them undissected) from Werribee, Port Phillip Bay, Victoria, Australia ( 37°57.085’ S, 144°47.128’ E) and paratype male from Davies Reef, Queensland, Australia ( 18° 48.78' S, 147° 39.30' E), partially dissected, ethanol-preserved; dissected parts mounted on slides in glycerine, sealed with Entellan®. Date of collection: 21st May 1985 and 21st September 1989, respectively. Slides deposited in the collection of MTQ, Australia (cat. MTQ W34377, MTQ W34378, MTQ 34379, MTQ 34380, MTQ W34381, respectively).
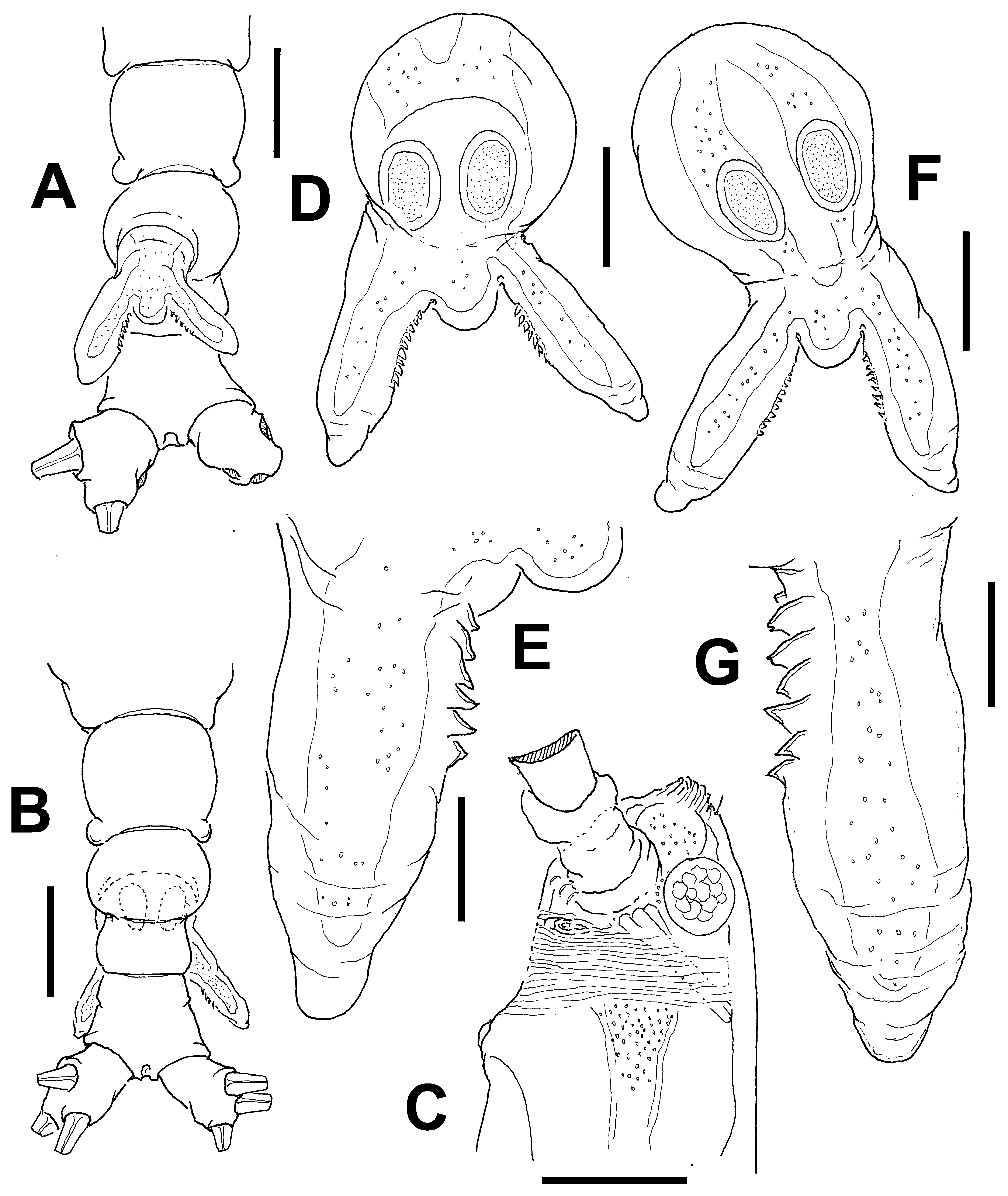
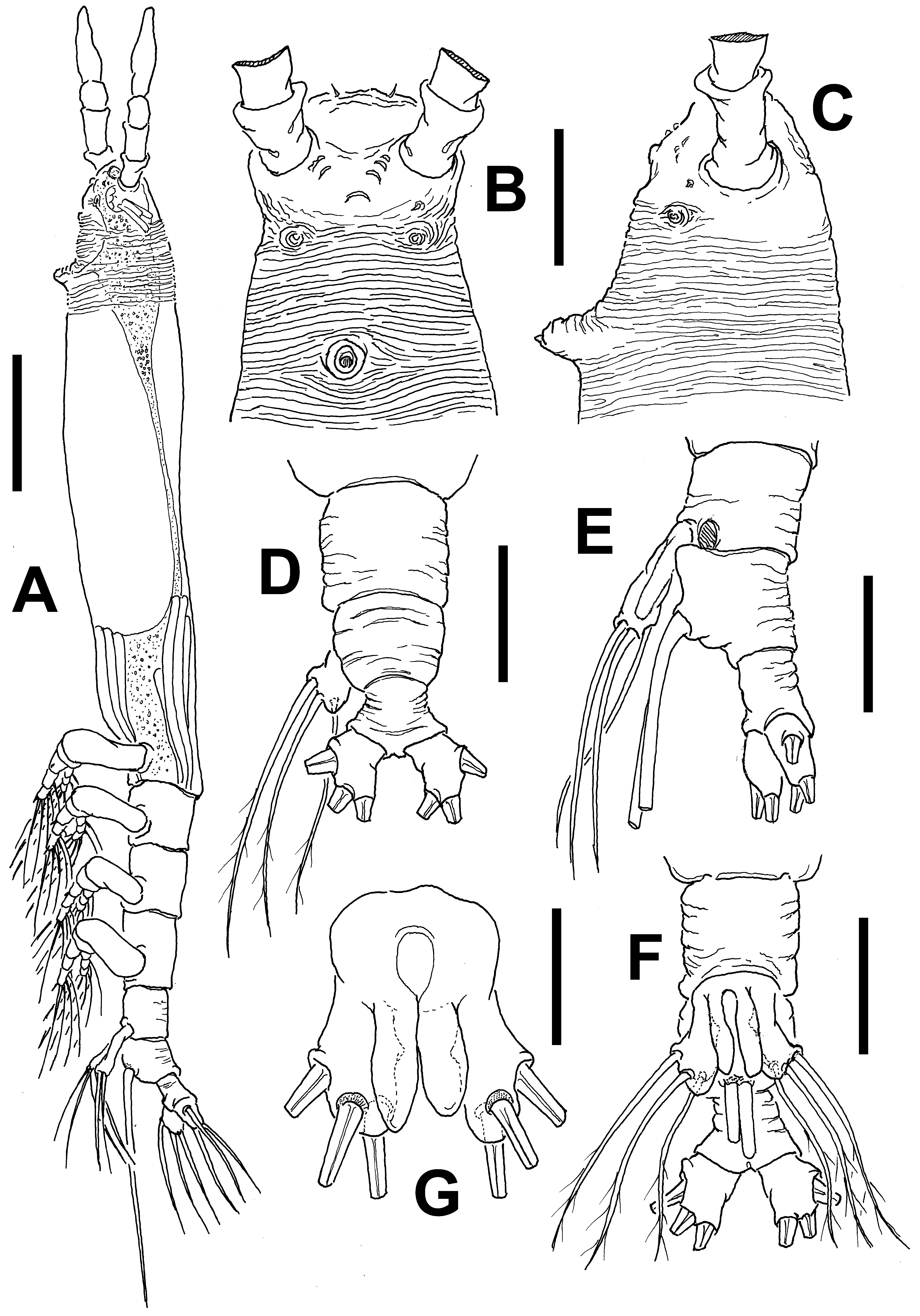
Description of adult female. Body noticeably elongate, slender ( Fig. 13 View FIGURE 13 A, D); body length of holotype female 1.65 mm. Cephalothorax approximately 1.0 mm long, representing 65% of total body length. Midventral oral papilla moderately protuberant, located at 20% of cephalothorax length. Pair of relatively large ocelli present, pigment cups moderately developed, medially conjoined, lightly pigmented; ventral cup larger than lateral cups ( Fig. 13 View FIGURE 13 D). Cephalic area laterally expanded, wider than cephalothorax. Frontal area ornamented with pattern of transverse striations ( Fig. 13 View FIGURE 13 C) with no frontal sensilla. Anterior 1/3 of cephalothorax ornamented with transverse striae arranged in tight regular ring-like pattern on ventral, lateral and dorsal surfaces ( Fig. 13 View FIGURE 13 A, B, C). Ventral surface with 1) pair of symmetrical, small nipple-like processes on anterior ventral surface located near bases of antennules, with no adjacent striae ( Fig. 13 View FIGURE 13 C); 2) second, larger nipple-like processes with concentric pattern of striae.
Urosome consisting of fifth pedigerous somite, genital double-somite and anal somite, together representing 15% of total body length. Relative lengths of urosomites (fifth pedigerous, genital double and free anal somites) 40.6: 31.2: 28.2 = 100, respectively ( Fig. 13 View FIGURE 13 E, D). Posterior half of fifth pedigerous somite with weak rounded lateral expansions. Genital double-somite longest of urosome, with smooth ventral and dorsal surfaces, lateral margins moderately swollen in dorsal view. Ovigerous spines paired, broken in examined specimen. Spines basally separated, slender, straight at their base ( Fig. 13 View FIGURE 13 A). Anal somite without medial constriction. Caudal ramus subquadrate, about as long as wide, armed with three caudal setae, broken off from proximal part in specimen examined ( Fig. 13 View FIGURE 13 F).
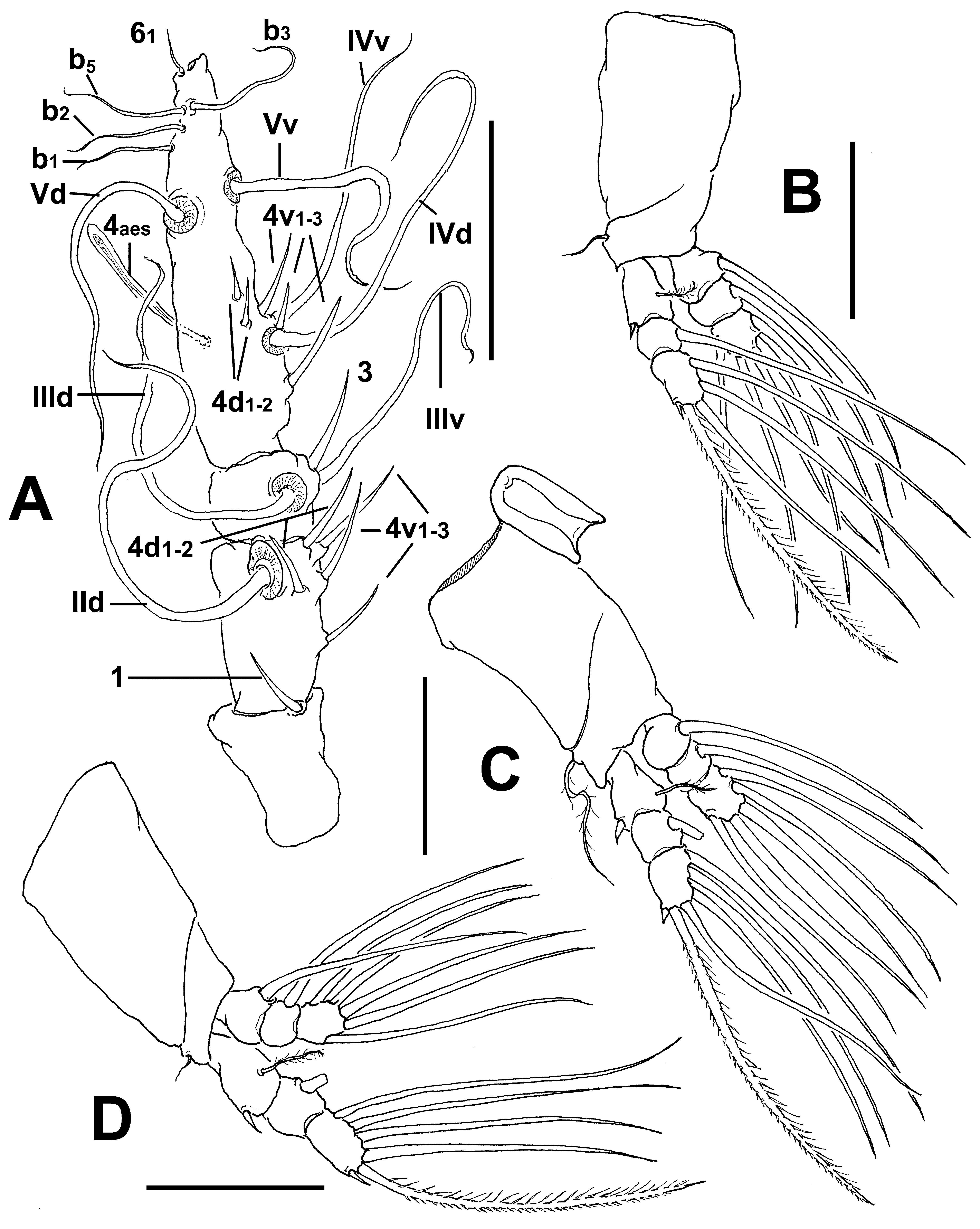
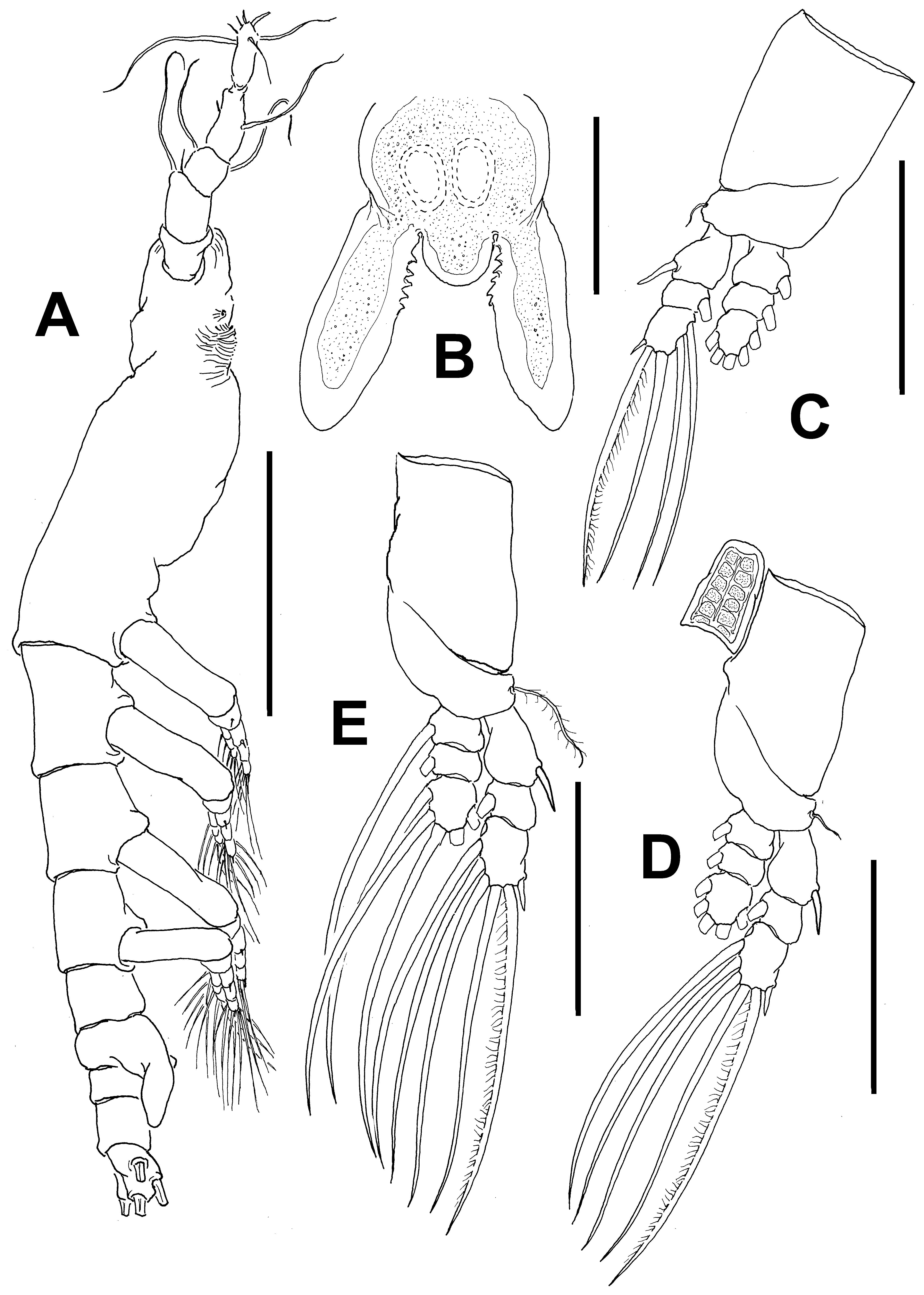
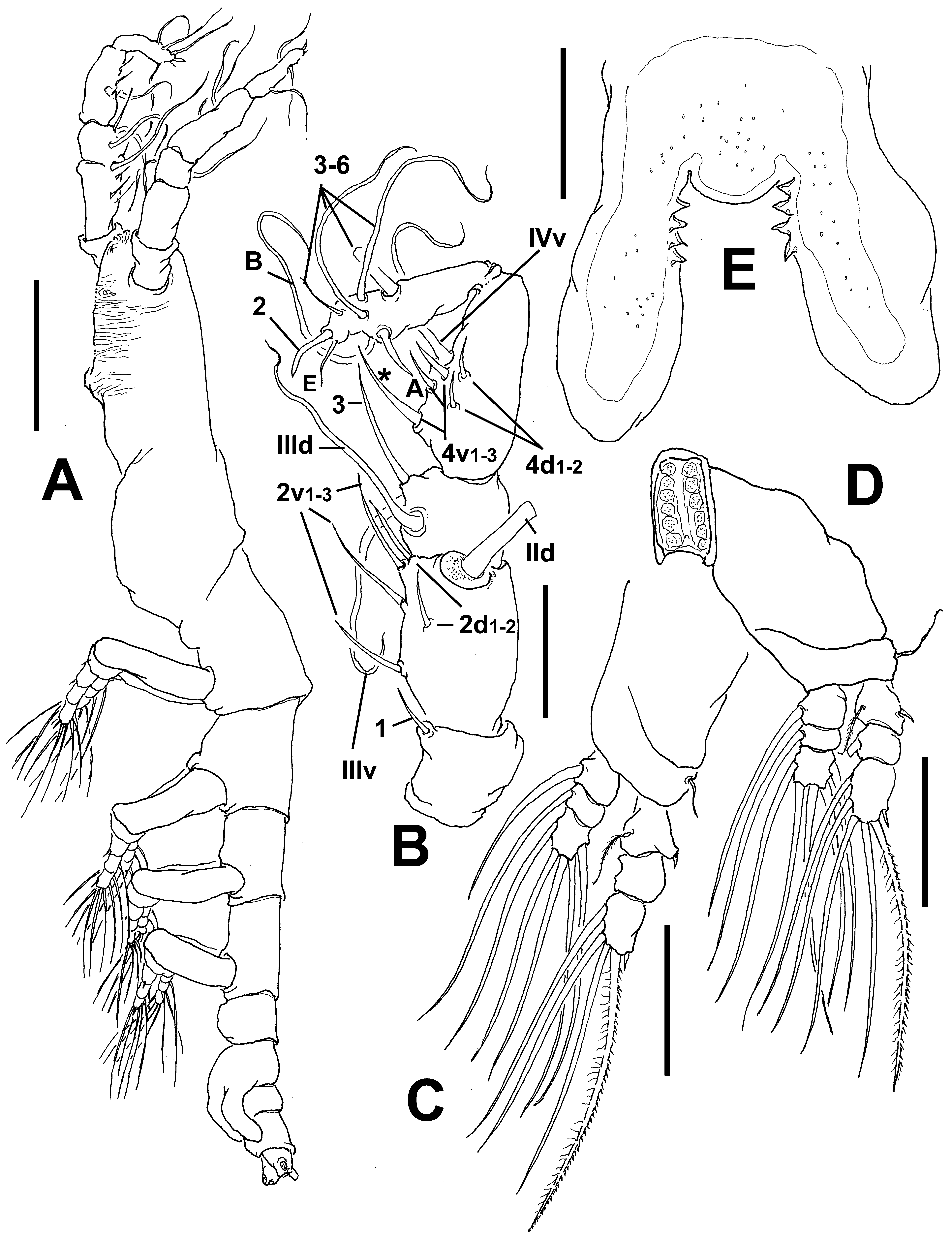
Antennule length 0.36 mm, representing about 22% of total body length and 34% of cephalothorax length; 4- segmented. Relative length of distal antennulary segment 52.3%. In terms of pattern described by Grygier & Ohtsuka (1995) for female monstrilloid antennulary armature, short, spiniform element 1 present on first segment; elements on second segment: 2d1-2, 2v 1-3, and IId. Third segment with element 3 being long, spiniform, elements IIId and IIIv setiform, of normal aspect. Segment 4 bearing elements 4d1,2, 4v 1-3, elements 4v 1,3 being longest of group; setae IVd, IVv, Vd, Vv, and element 4aes present. Subterminal elements b1-3, 6 present, unbranched; element 61 present, 62 absent in specimen, but socket was observed ( Fig. 14 View FIGURE 14 A).
Incorporated first pedigerous somite and succeeding three free pedigerous somites each bearing a pair of biramous legs. Pedigerous somites 2–4, together accounting for 22% of total body length. Intercoxal sclerites of legs 1–4 subrectangular, surface and posterior margin smooth ( Fig. 14 View FIGURE 14 C). Bases of legs articulating with large, rectangular coxae along oblique line; with hair-like lateral seta ( Figs. 14 View FIGURE 14 B–D); on leg 3, this seta about five times longer, sparsely setulated and thicker than those on other legs ( Fig. 14 View FIGURE 14 C). Endopods and exopods of legs 1–4 triarticulated. Ramal setae all biserially plumose except spiniform outer seta on exopodal segments1 and 3, and inner seta of first exopodal segment, these latter being short, slender, and sparsely setulated. Outermost distal spines on third exopodal segment of legs 1–4 short, 0.25 times as long as segment. Outermost apical exopodal setae of legs 1–4 with inner margin setulose, outer margin spinulose.
Armature formula of legs 1–4: Fifth legs medially conjoined, bilobate, outer (exopodal) lobe distally expanded. Outer lobe armed with three subdistal setae ( Fig. 13 View FIGURE 13 E, F). Inner lobe with similar digitiform shape, shorter than outer lobe, unarmed, reaching about ¾ the length of outer lobe.
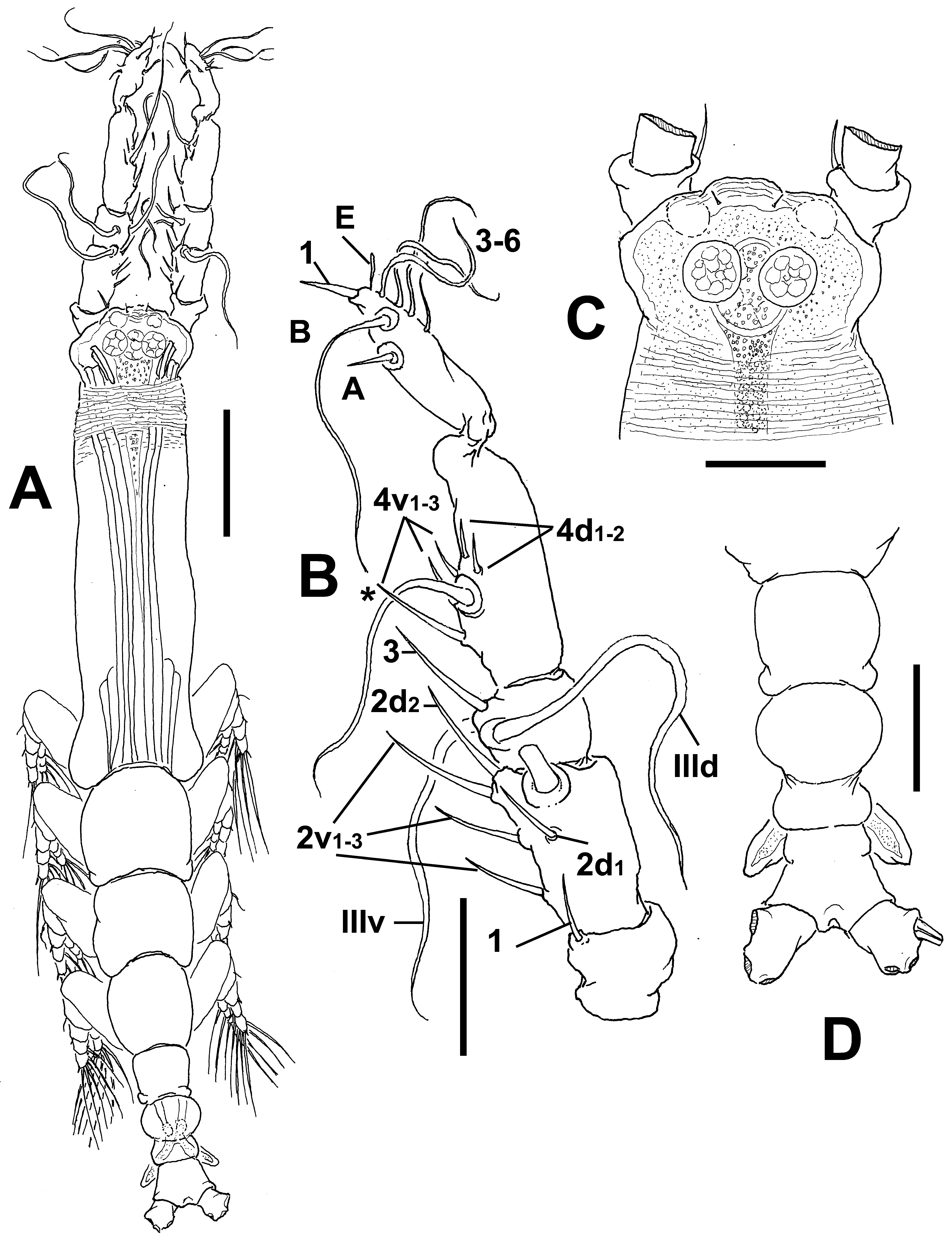
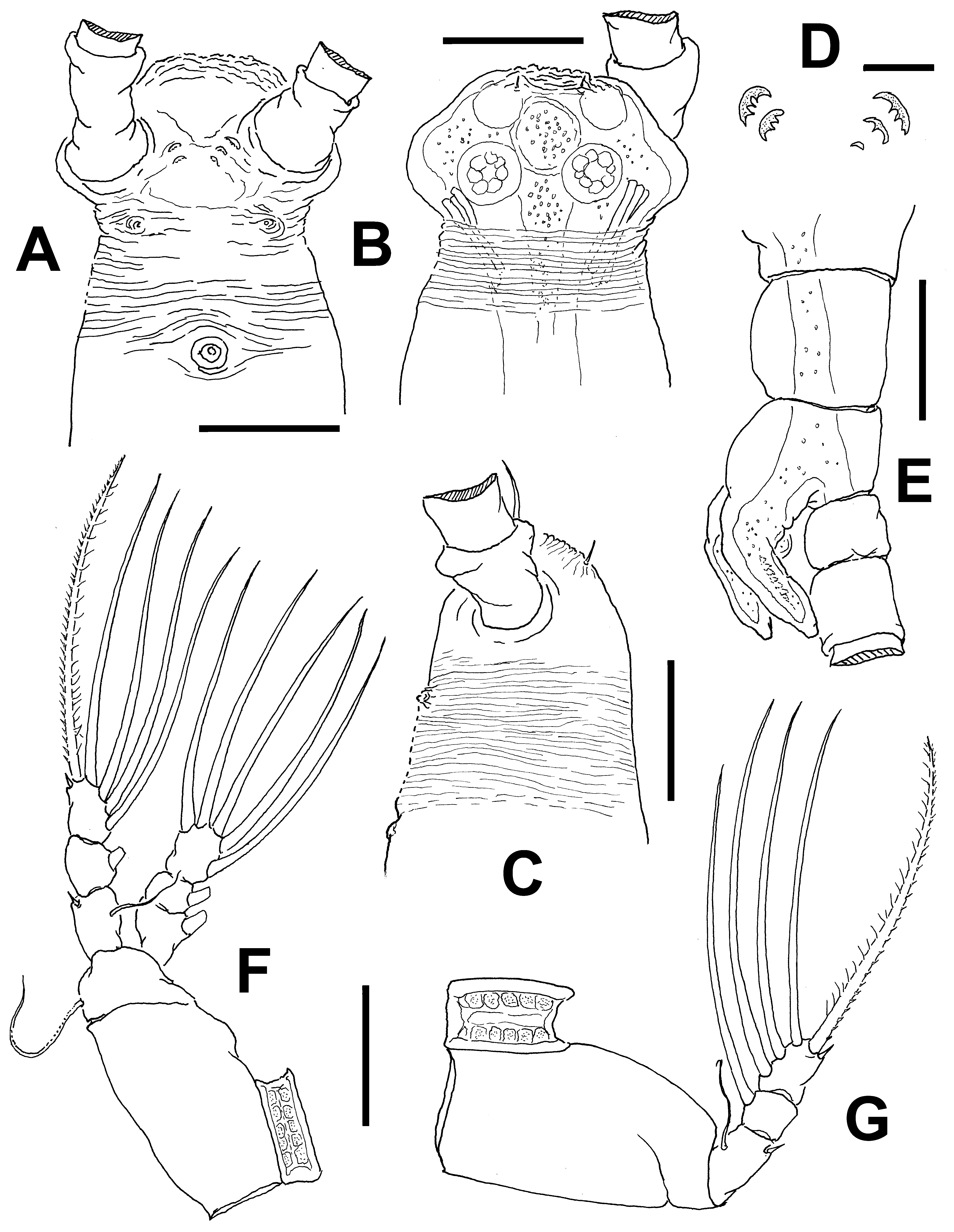
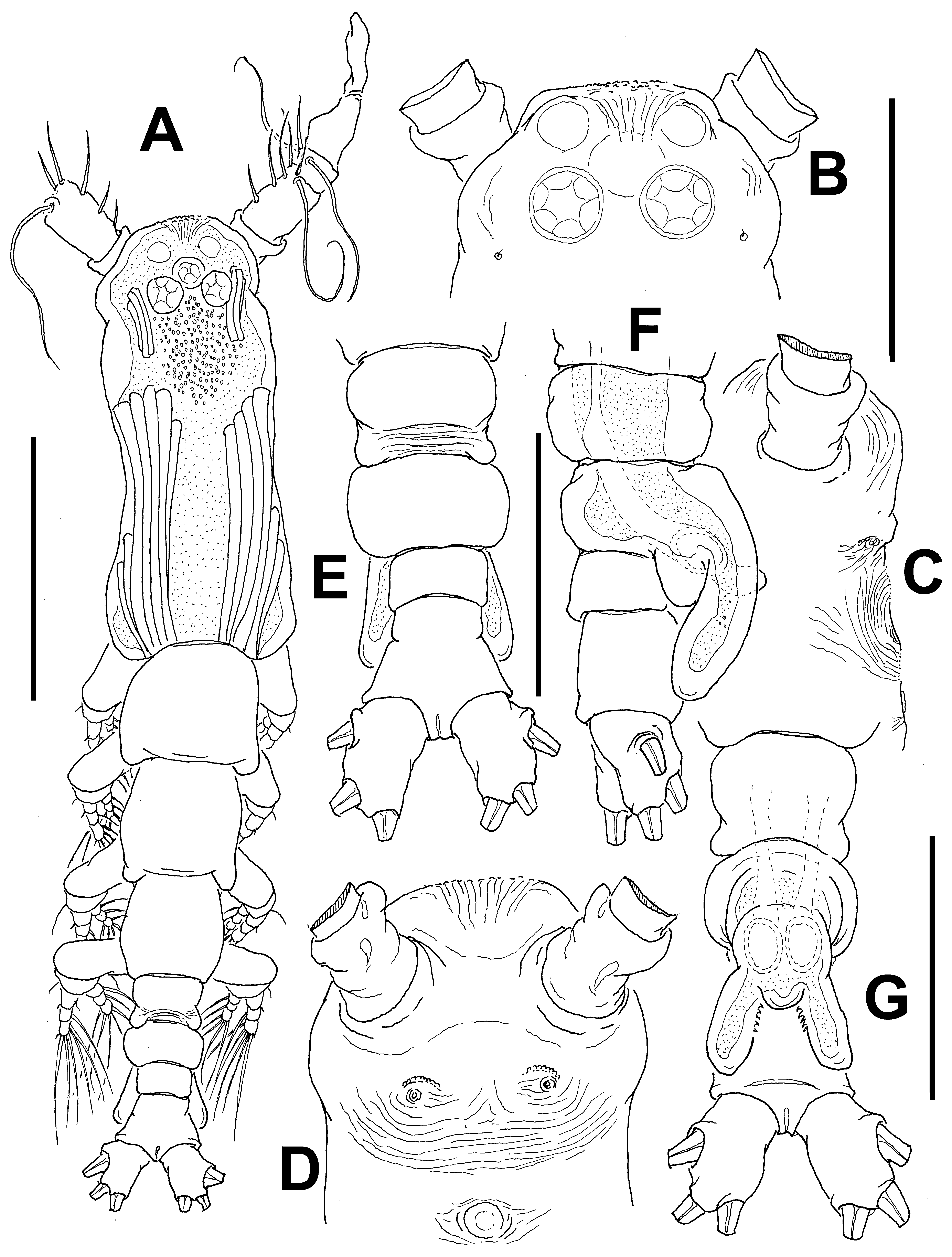
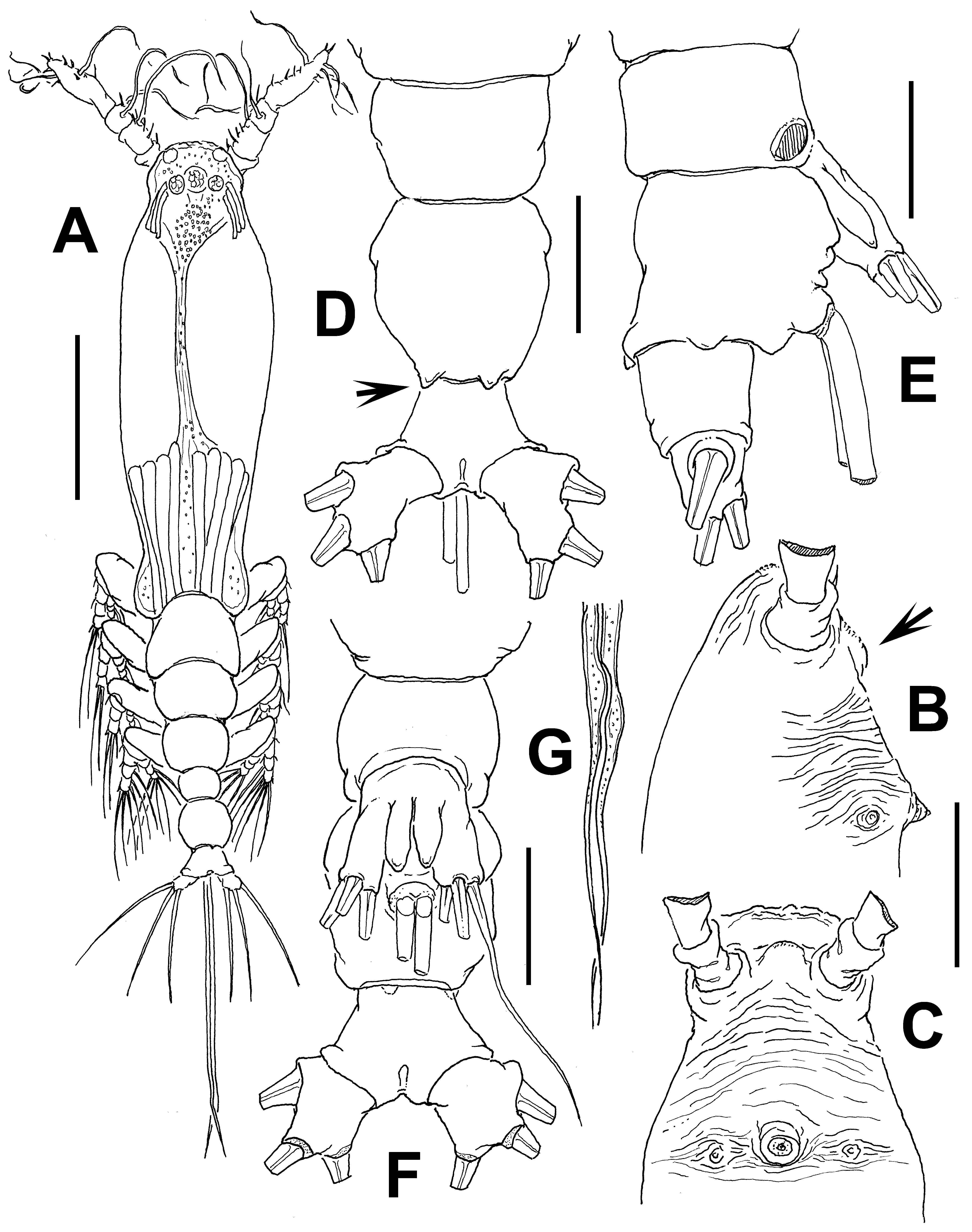
Description of male. Total body length of allotype male from Werribee = 1.24 mm. Size range of male specimens = 0.64 (Davies Reef) – 1.44 mm (Werribee); average ( n =4) 1.23 mm. Cephalothorax 0.34–0.73 mm long, representing 51% of total body length ( Figs 15 View FIGURE 15 A, 18A, 19A). Midventral oral papilla weakly developed, located at 24% of cephalothorax length. Cephalic region moderately ( Fig. 15 View FIGURE 15 A) or weakly ( Fig. 18 View FIGURE 18 A) protuberant bilaterally in dorsal view, with fringe of striae surrounding the cephalic region as in female (males of Werribee) or restricted to ventral area ( Fig. 18 View FIGURE 18 D), in both groups limited to anterior 1/5 of cephalothorax ( Figs. 16 View FIGURE 16 A–C). Pair of dorsal ocelli present, weakly developed; pigment cups medium-sized. Ocelli separated by the length of less than one eye diameter, faintly pigmented. Pair of sensilla between antennulary bases present (Werribee) or absent (Davies Reef). Forehead area as in female, ornamented with transverse ( Fig. 16 View FIGURE 16 A, B) or parallel ( Fig. 18 View FIGURE 18 B) cuticular ridges. Other cuticular processes include: 1) three pairs of cuticular crescent-shaped processes between antennule bases ( Fig. 16 View FIGURE 16 D), absent in specimen from Davies Reef ( Fig. 18 View FIGURE 18 D); 2) nipple-like processes surrounded by concentric and transverse striae.
Urosome consisting of fifth pedigerous, genital somite (carrying genital complex), preanal somite, and anal somite. Fifth pedigerous somite with smooth ventral surface, with rounded postero-lateral processes as in female in both specimens from Werribee ( Figs 15 View FIGURE 15 D, 17B) and Davies Reef, which has transverse wrinkles in distal half of somite ( Fig. 18 View FIGURE 18 E). Genital somite slightly shorter than fifth pedigerous somite. Genital somite with smooth dorsal surface and expanded lateral margins. Preanal somite short, smooth, without constriction. Genital complex of type II ( Suárez-Morales & McKinnon 2014), represented by pair of moderately divergent, digitiform genital lappets. Lappets reach beyond midlength of anal somite, ornamented with row of 6–10 sharp dentiform processes along inner margins ( Figs 17 View FIGURE 17 A, B, D–G; 18E, F, G; 19B). Rounded, protuberant medial process present at common basal joint of lappets ( Figs. 17 View FIGURE 17 , 18 View FIGURE 18 ). Anal somite about twice as long as preanal somite in dorsal and lateral views, comprising 26% of urosome length; weak lateral constriction visible in dorsal aspect ( Fig 17 View FIGURE 17 B, 18E). Caudal rami subquadrate, approximately 1.1 times as long as wide, about as long as anal somite. Each ramus with three setae.
Antennulary length 0.49 mm (male from Werribee) and 0.21 mm (male from Davies Reef). Antennules relatively short, representing 34% of total body length, and 66% of cephalothorax length; 5-segmented, all segments separated, with segment 5 located distal to geniculation ( Figs. 15 View FIGURE 15 B, 18A). Setal element 1 on first segment slender, spiniform, moderately long, reaching proximal 1/3 of second segment. Antennulary elements 2v 1-3, 2d1,2 long, slender as in female; element IId present on second segment. Setal elements IIId, IIIv, and long element 3 present on third segment. Fourth segment with elements 4d1,2, 4v 1–3 present; element 4v 3 being longest of group. Fifth segment with 4 “b”-group setae, elements b1-3 slender, unbranched; element 61 present in distal position. According to Huys et al. (2007) setal nomenclature of distal segment, elements A, B, and E and 1, 3–6 present ( Fig. 15 View FIGURE 15 B).
Incorporated first pedigerous somite and succeeding three pedigerous somites each bearing well-developed biramous legs. Pedigerous somites 2–4, together accounting for 31% of total body length in dorsal view. Legs 1–4 as in female except for surface of coxal plates ornamented with spinulose patches ( Figs 16 View FIGURE 16 F, G, 19 C–D).
Type locality. Werribee, Port Phillip Bay, Victoria, Australia ( 37°57.085’ S, 144°47.128’ E).
Etymology. The species name, an adjective in apposition, results from the combination of two Latin terms, annulus (= ring) and collum (= neck); the name refers to the appearance of the post-cephalic (“neck”) region of the cephalothorax, with deep transverse, parallel ring-like striae.
Diagnosis. Cymbasoma with long cephalothorax, representing 65% of total body length, third antennulary segment representing more than 52% of antennule length, with two pairs of nipple- like processes on ventral surface of cephalic region. Cephalothorax with extense, complete fringe of striation covering about 1/3 of cephalothorax. Genital double-somite with lateral margins expanded and with rounded postero-lateral processes.
Anal somite without medial constriction. Fifth leg with elongate, thumb-like outer lobe with three subdistal setae, inner lobe also thumb- like, smaller than outer lobe, unarmed. Male with relatively reduced fringe of striae around cephalic region. Genital complex of Type II; genital lappets ornamented with 6–10 sharp dentiform processes along inner margins and with rounded process at common basal joint of lappets.
Remarks. The female of this species can be distinguished by its possession of a wide fringe of cuticular striae covering about 1/3 of the cephalothorax. The striae are weaker posteriorly but cover all around the anterior 40% of the cephalothorax ( Fig. 13 View FIGURE 13 B). The cephalothorax is elongate, representing up to 65% of the total body length, a character shared with species of the C. longispinosum -group, but differs in the structure of the ovigerous spines, which are basally separated in the new species ( Fig. 13 View FIGURE 13 E) but proximally fused in species of the C. longispinosum - group ( Üstün et al. 2014). A similar pattern of cephalic striation is present in C. striifrons ( cf. Chang 2012) and also in the Australian C. markhasevae sp. nov. ( Fig. 32 View FIGURE 32 A–C), but the body is different in these species. In C. striifrons it is clearly shorter and more robust than in the new species. In C. markhasevae sp. nov. the body proportions are similar to C. annulocolle but the fifth pedigerous somite and the genital double-somite differ in these species; in the former the fifth pedigerous somite has straight lateral margins and lacks the postero-lateral rounded processes present in the latter. The genital double-somite is clearly wider and shorter in C. markhasevae sp. nov. than in the new species; this somite is slightly longer than the fifth pedigerous somite whereas it is clearly shorter in C. markhasevae ( Fig. 32 View FIGURE 32 D). The antero-ventral process of the genital double-somite is weakly developed in the new species but it is clearly stronger in C. markhasevae ( Fig. 32 View FIGURE 32 E).
In addition, the coverage and width of the striation area is clearly more extensive in the new species than in C. striifrons , which has a narrow fringe ( Chang 2012) and in C. markhasevae , in which the fringe ends abruptly at the post-oral area and no posterior fainter wrinkles are discernible as in the new species. Also, the armature of the fifth leg outer (exopodal) lobe of the new species is distinctive for its long innermost seta, which is the longest of the three; this is a character not observed in other similar species in which the innermost seta is usually shorter or as long as the other two on this lobe.
Another species of Cymbasoma with a similar striation pattern stretching around the cephalothorax is C. striatus ( Isaac, 1974) ( Suárez-Morales 2000b) but in contrast to the new species its fifth leg has only one lobe, the endopodal lobe being absent. The new species shares with the Australian C. bidentatum sp. nov. ( Fig. 20 View FIGURE 20 F) and C. markhasevae ( Fig. 32 View FIGURE 32 G) an inner lobe of the fifth leg that is elongate, narrow, and arising proximally from the same base as the outer lobe. These species differ in several other features as explained in the respective comparative sections below.
The male of this species can be distinguished from its known congeners by the presence of a row of 6–10 spines along the inner margin of the genital lappets. Such ornamentation is present only in C. quadridens in which the spines are smaller and fewer (4–6) and the lappets are relatively shorter and wider than in the new species ( Suárez-Morales & Pilz 2008: fig. 7B, C). In addition, the new species ( C. annulocolle ) has a wrinkled frontal surface vs. a smooth surface in C. quadridens ( Suárez-Morales & Pilz 2008: figs 5D, 7A). The Australian C. pseudoquadridens sp. nov. has also spines on the lappets but these are clearly larger and stronger and only up to four in number ( Fig. 23 View FIGURE 23 E), thus diverging from the pattern observed in the new species. Also, the fringe of striae present in the new species is absent from both C. quadridens ( cf. Davis 1947; Suárez-Morales & Pilz 2008: fig. 5A) and C. pseudoquadridens ( Fig. 32 View FIGURE 32 A). The new species differs from the closely related C. quadridens or C. tenue by having an inner serra-like ornamentation on each lappet, with 6–10 dentiform processes vs. only one proximal spiniform process in C. tenue ; differences with respect to C. quadridens ( Davis 1947; Suárez-Morales & Pilz 2008) were discussed above.
One of the criteria to link males and females of a species is to find them in the same sample/locality, which is partially the case for the male and the female of this species, but a male was found in a different area. A second factor is to evaluate if they share non-sexually related characters; in C. annulocolle both sexes have 1) a distinctive ornamentation of the cephalothorax, with a band of striae, and 2) a pair of rounded posterolateral process on the fifth pedigerous somite. These two characters are not present in other species in the same sample/area. Furthermore, they also share a similar ornamentation pattern of antennulary segments 1–4. So, these comparative and distributional analyses lead us to match both genders instead of erecting a new taxon for the males of this species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |