
Panonychus caricae Hatzinikolis, 1984
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4881.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:0F7AD901-BE3E-4799-98A3-0F8DF83E3818 |
|
DOI |
https://doi.org/10.5281/zenodo.4328182 |
|
persistent identifier |
https://treatment.plazi.org/id/03C4F678-FFDF-FFB0-0BFC-F3C8F855F9CE |
|
treatment provided by |
Plazi |
|
scientific name |
Panonychus caricae Hatzinikolis, 1984 |
| status |
|
Panonychus caricae Hatzinikolis, 1984
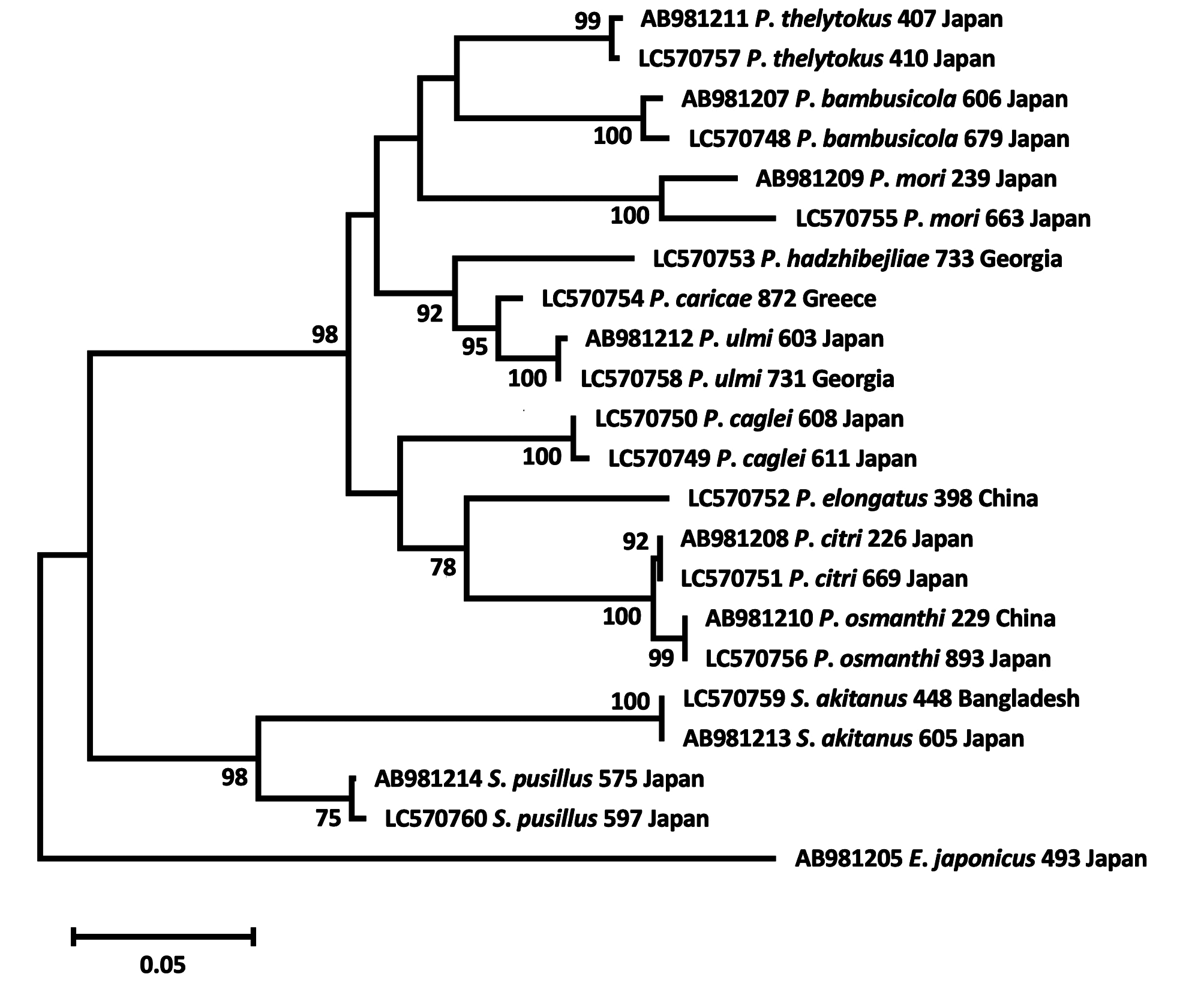
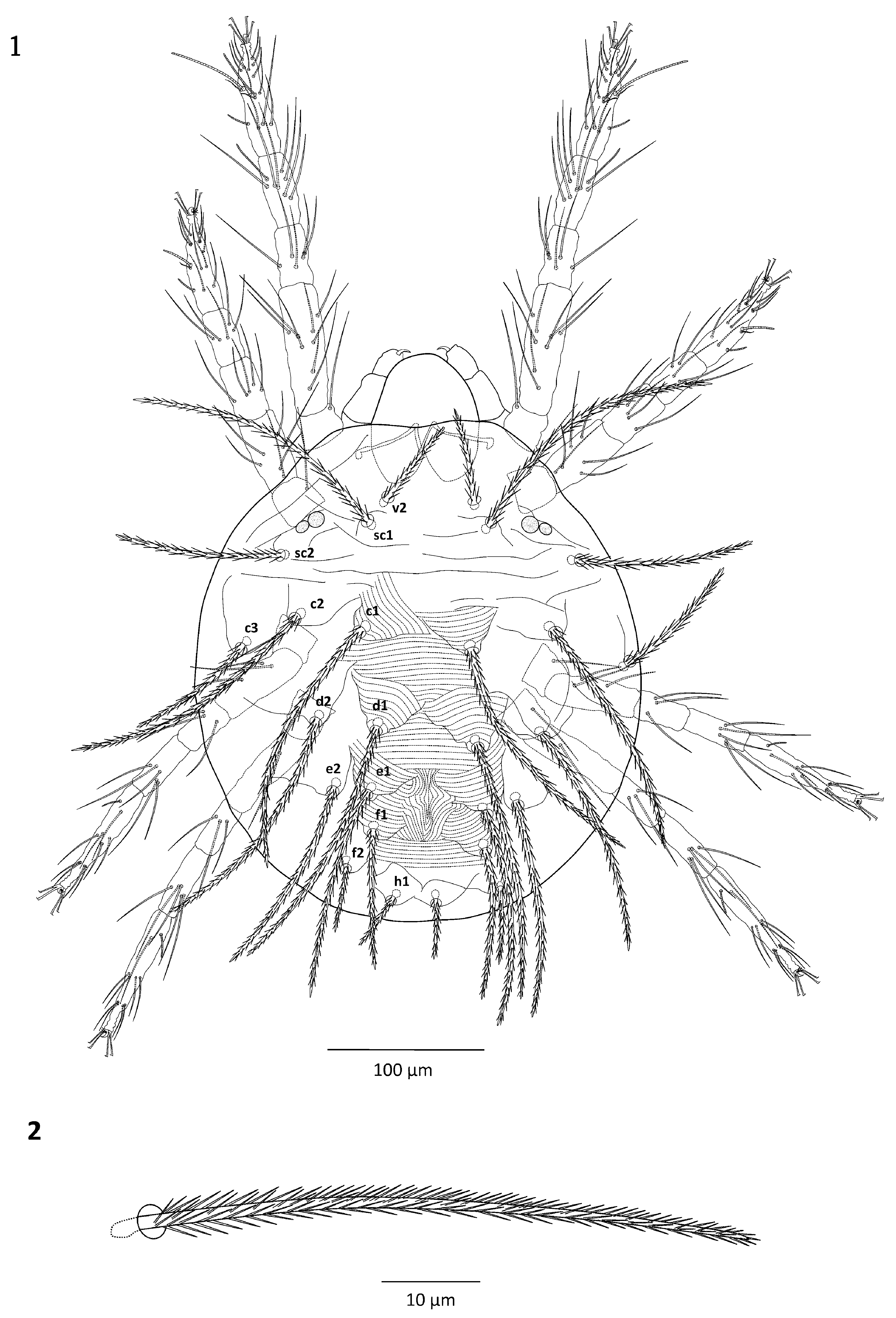
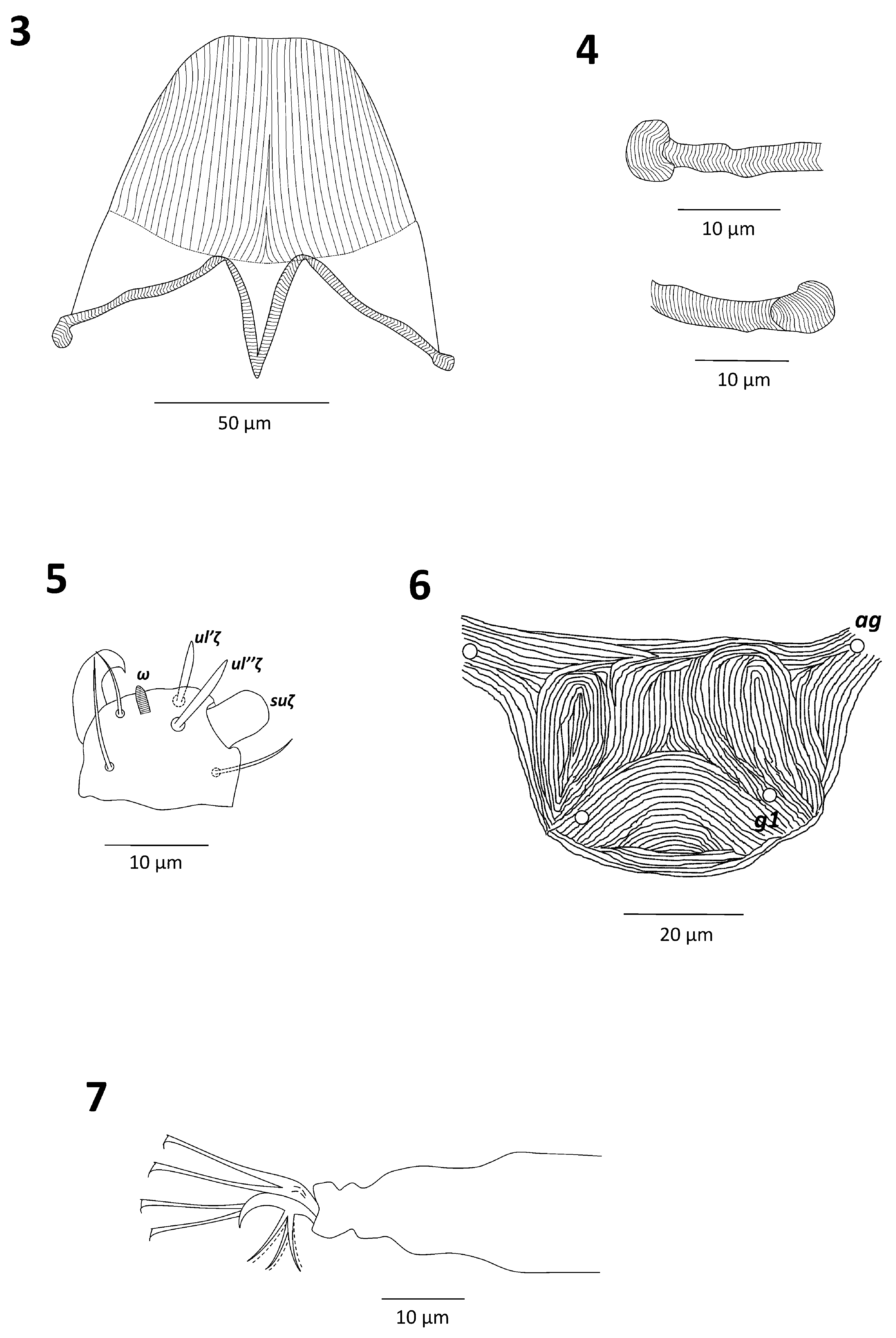
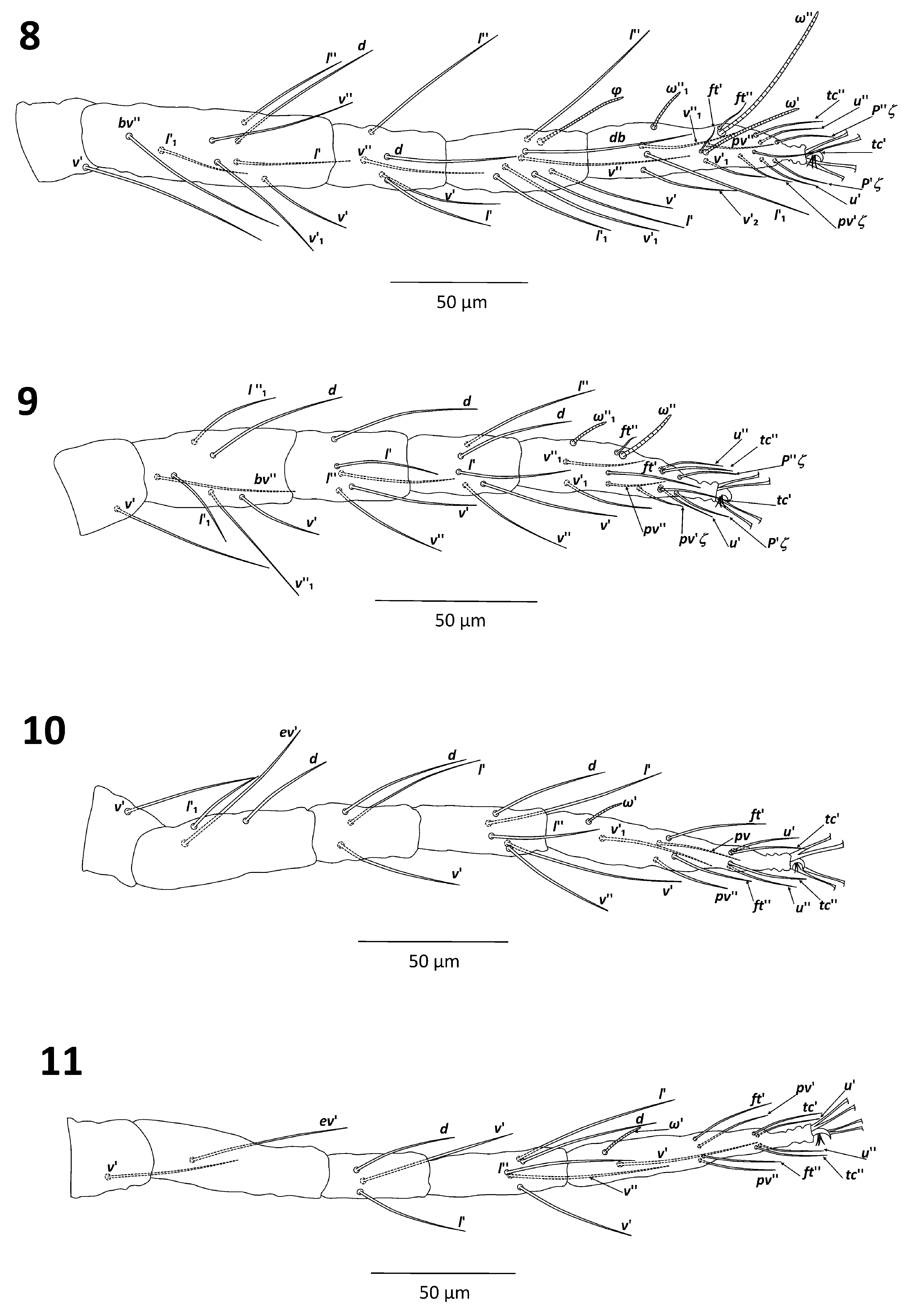
( Figs. 1–21 View FIGURES 1–2 View FIGURES 3–7 View FIGURES 8–11 View FIGURES 12–15 View FIGURES 16–21 )
Diagnosis. Dorsal setae stout, lanceolate, coarsely barbed, much longer than longitudinal intervals between setal rows, set on well-developed white tubercles; striation pattern on genital flap transverse, arched; pregenital striae longitudinal-oblique, becoming longitudinal immediately anterior to genital flap. Peritreme straight with short distal bulb; distal bulb sometimes forming short hook resembling a golf-club. Tarsus I of female with 3 tactile setae and 1 solenidion proximal to duplex setae; tibiae I of both sexes with 7 tactile setae; tibiae II, III and IV of both sexes with 5 tactile setae; femora IV of both sexes with 1 seta. Empodia I–IV split into 3 pairs of medioventral hairs. Aedeagus directed dorsally, shaft weakly sigmoid and weakly tapering to blunt tip.
Re-description. Female (n=10; Figs. 1–11 View FIGURES 1–2 View FIGURES 3–7 View FIGURES 8–11 ). Body oval and reddish. Length of body including gnathosoma 330–430; width of body in humeral part 250–310.
Dorsum. Dorsum with 13 pairs of long, coarsely barbed setae ( Figs. 1–2 View FIGURES 1–2 ); Opisthosomal setae set on strong white tubercles; length of setae: v 2 50–62; sc 1 155–175; sc 2 85–112; c 1 155–185; c 2 145–175; c 3 70–92; d 1 155–187; d 2 125–160; e 1 137–162; e 2 100–132; f 1 80–102; f 2 42–50; h 1 35–40 ( Fig. 1 View FIGURES 1–2 ).
Distances between bases of body setae: v 2 – v 2 50–60; sc 1 – sc 1 67–87; sc 2 – sc 2 175–205; c 1 – c 1 50–75; c 2 – c 2 150180; c 3 – c 3 230–280; d 1 – d 1 50–95; d 2 – d 2 142–170; e 1 – e 1 55 –77; e 2 – e 2 117–150; f 1 – f 1 60–77; f 2 – f 2 80–120; h 1 – h 1 27–37; v 2 – h 1 220–300; c 1 – d 1 50–70; d 1 – e 1 37 –65; e 1 – f 1 25–37; f 1 – h 1 27–52.
Dorsocentral striae of the opisthosoma transverse between setae c 1 and d 1, and d 1 and e 1; irregularly longitudinal between setae e 1 and f 1, forming a diamond-shape pattern ( Fig. 1 View FIGURES 1–2 ).
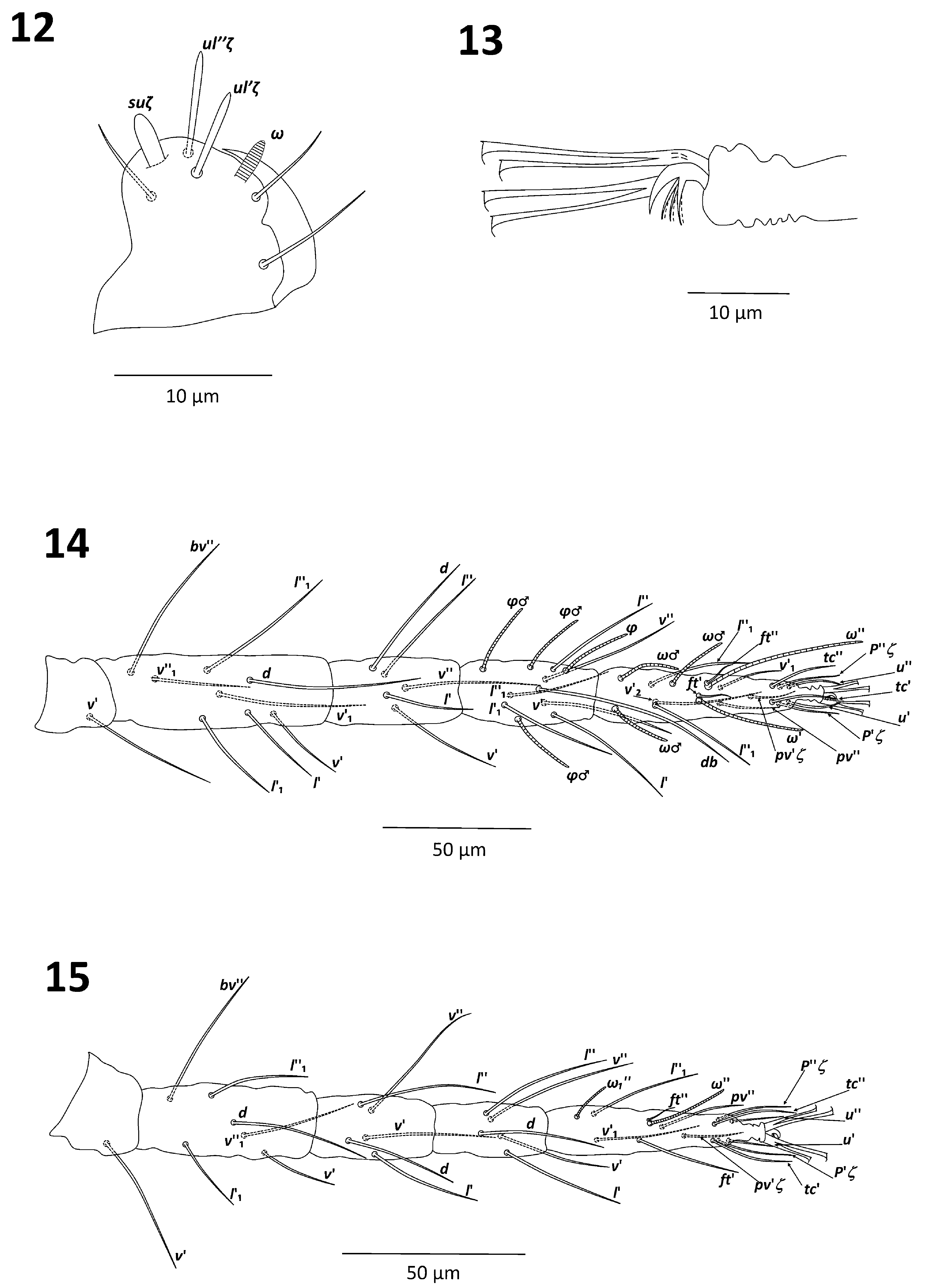
Gnathosoma. Stylophore rounded anteriorly with longitudinal, parallel striation ( Fig. 3 View FIGURES 3–7 ). Peritremes simple, ending with short distal bulb; distal bulb sometimes forming short hook resembling a golf-club ( Figs. 3–4 View FIGURES 3–7 ). Terminal sensillum suζ (spinneret) of palptarsus broad, approximately 1.5 times as long as wide: length 5–7 and width 4–5; dorsal sensillum (solenidion, ω) shorter than spinneret 3–5 long; eupathidia different in length: ul”ζ longer than ul’ζ, 8–10 and 6–7 long, respectively ( Fig. 5 View FIGURES 3–7 ).
Venter. Striae on genital flap entirely transverse, arched in the middle part of the flap; pregenital area with longitudinal striae anterior to genital flap, striae varying from longitudinal to oblique laterally ( Fig. 6 View FIGURES 3–7 ); all ventral setae smooth and thin.
Legs. Leg I 270–300, leg II 200–240, leg III 210–260; leg IV 230–280 long (from trochanter to tarsus). Length of segments of leg I as follows: femur 75–87.5, genu 37–45, tibia 42–50, tarsus 92–97 long. Leg setal counts as follows ( Figs. 8–11 View FIGURES 8–11 ):
I 2–1–8–5–7 + (1)–12 + (1) + 2 duplexes
II 2–1–6–5–5–11 + (1) + 1 duplex
III 1–1–3–3–5–9 + (1)
IV 1–1–1–3–5–9 + (1)
Tarsus I with 2 duplex setae, distal duplex solenidion (ω”) of tarsus I more than two times longer than proximal duplex solenidion (ω’): 57–63 and 22–30 long, respectively; tactile setae (ft’, ft”) of distal and proximal duplexes subequal in length: 10–12; tarsus I with 3 tactile setae and 1 solenidion proximal to proximal set of duplex setae ( Fig. 8 View FIGURES 8–11 ). Tarsus II with 1 duplex seta, solenidion of duplex seta 20–25 long and tactile seta 10–12 long; tarsus II with 2 tactile setae and 1 solenidion proximal to duplex seta, other 2 tactile setae near the level of duplex seta; tibia II with 5 tactile setae ( Fig. 9 View FIGURES 8–11 ). Tibiae III and IV with 5 tactile setae ( Figs. 10–11 View FIGURES 8–11 ).
Empodia I–IV split into two sets of three ventrally directed hairs of uniform length ( Figs. 7–11 View FIGURES 3–7 View FIGURES 8–11 ).
Male (n=10; Figs. 12–21 View FIGURES 12–15 View FIGURES 16–21 ). Reddish green to pale red in color. Length of body including gnathosoma 310–350; width of body 170–190.
Dorsum. Dorsal body setae stout, strong, longer than distance between bases of consecutive setae, length of setae: v 2 45–55; sc 1 73–122; sc 2 65–95; c 1 85–125; c 2 80–120; c 3 60–87; d 1 78–117; d 2 75–115; e 1 50 –62; e 2 50 –72; f 1 30–40; f 2 25–30; h 1 15–22.
Distances between bases of the body setae: v 2 – v 2 40–52; sc 1 – sc 1 55–65; sc 2 – sc 2 125–152; c 1 – c 1 37–57; c 2 – c 2 87–112; c 3 – c 3 137–165; d 1 – d 1 37–52; d 2 – d 2 92–130; e 1 – e 1 32 –37; e 2 – e 2 67–75; f 1 – f 1 27–35; f 2 – f 2 45–50; h 1 – h 1 15–25; v 2 – h 1 175–200; c 1 – d 1 30–37; d 1 – e 1 30 –37; e 1 – f 1 25–30; f 1 – h 1 17–22.
Gnathosoma. Stylophore and peritreme as in female ( Figs. 3–4 View FIGURES 3–7 ). Terminal sensillum suζ (spinneret) of palptarsus approximately two times as long as wide: 4–5 long and 2 wide; dorsal sensillum (solenidion, ω) 3–4 long, slightly shorter than spinneret; eupathidia ul”ζ longer than ul’ζ, 7–9 and 5–7 long, respectively ( Fig. 12 View FIGURES 12–15 ).
Legs. Leg I 280–330; leg II 210–250; leg III 230–300; leg IV 240–300 long (from trochanter to tarsus). Length of first leg segments as follows: femur 77–100; genu 42–52; tibia 47–57; tarsus 95–112 long. Leg setal count as follows ( Figs 14–17 View FIGURES 12–15 View FIGURES 16–21 ):
I 2–1–8–5–7 + (4)–12 + (3) + 2 duplexes
II 2–1–6–5–5–11 + (1) + 1 duplex
III 1–1–3–3–5–9 + (1)
IV 1–1–1–3–5–9 + (1)
Tarsus I with 3 tactile setae and 3 solenidia proximal to proximal set of duplex setae; distal duplex solenidion of tarsus I longer than that of proximal duplex: 57–65 and 32–47 long, respectively; tactile seta of distal and proximal duplexes equal in length: 12–15 ( Fig. 14 View FIGURES 12–15 ). Tarsus II with 2 tactile setae and 1 solenidion proximal to duplex seta, one tactile seta near level of duplex seta; tarsus II solenidion of duplex seta 22–30 long, tactile 12–15 long ( Fig. 15 View FIGURES 12–15 ). Tibiae II, III and IV each with 5 tactile setae ( Figs. 15–17 View FIGURES 12–15 View FIGURES 16–21 ).
Empodia I–IV as female split into two sets of three ventrally directed hairs of uniform length ( Fig. 13 View FIGURES 12–15 ).
Aedeagus. Aedeagus dorsally directed, shaft bents nearly at right angle, ending with weakly sigmoid blunt tip; bent portion of aedeagus shorter than dorsal margin of shaft ( Figs. 18–21 View FIGURES 16–21 ). Measurements of aedeagus: shaft 11–13 length, width (broadest part of shaft) 5–6; bent portion of aedeagus 6–8 length.
Specimens examined. 10 females and 10 males; ex. leaves of Ficus sp. ( Moraceae ), Ano Drosia , Kalamata, Greece (37°02’18’’N, 21°51’17’’E); 9 September 2017; leg. T. Gotoh .
Specimens depository. 6 females and 6 males are deposited in the Institute of Zoology , Ilia State University, Georgia ; 10 females and 17 males are deposited in the Laboratory of Applied Entomology and Zoology, Ibaraki University ( AEZIU) with the voucher specimen no. 872 .
Remarks. Unfortunately, our attempts to find the type specimens were unsuccessful, so the only possible comparisons are with the original description. Hatzinikolis (1984) measured the holotype only, making statistical comparison—such as that of Arabuli et al. (2016) —impossible. However, both species were found in Greece, the host plant species is the same, the general morphology is similar, and the setal lengths are within the range of our specimens, or very close to them (e.g. f 2 44 µm and h 1 42 in the holotype description versus 42–50 and 35–40, respectively). Therefore, we have tentatively classified our specimens as P. caricae .
Some differences seem to be apparent in the palp-tarsus, with the solenidion drawn as approximately the same length of the spinneret, and the spinneret measurements of 11 x 9 μm. However, without type specimens, caution is needed interpreting the drawing of Hatzinikolis (1984), especially as the scale and measurements are probably incorrect. According to the drawing, the palp length (ca. 220 μm) is greater than the body width (198 μm), which must be wrong, and it seems more likely that the scale is approximately double (i.e. 5 μm, not 10 μm). If so, the spinneret measurements are approximately the same as in our specimens.
The re-described species can be separated from other Panonychus species using the length of the female dorsal setae in combination with the ratio between the length of female dorsal opisthosomal setae f 2 and h 1, and the ratio between the length of dorsal setae sc 1 and h 1. According to our morphological examinations, P. caricae is closely related to P. ulmi and P. hadzhibejliae . These species have the similar chaetotaxy of legs, and the shape of aedeagus and the angle between the sigmoid distal part and ventral margin of the shaft are comparable. However, P. caricae can be easily distinguished from P. ulmi by the length of the dorsal opisthosomal setae: c 3 111 (100–117), e 2 147 (137–150), f 2 66 (62–70) for P. ulmi and 85 (70–92), 120 (100–132), 50 (42–50) for P. caricae .
The species P. hadzhibejliae and P. caricae seem more closely related morphologically and were collected on the same host plant species ( Arabuli et al. 2016). Panonychus caricae has longer setae sc 1 (155–157) and dorsocentral setae c 1 (155–185), c 2 (144–185), d 1 (115–150) and e 1 (137–162) compared with P. hadzhibejliae (sc 1 120–145, c 1 112–140, c 2 115–140, d 1 115–150, e 1 110–135) and, in general, most other dorsal setae of P. caricae are in the upper limits or longer than seen in P. hadzhibejliae , with the exception of f 2 and h 1. Furthermore, our molecular analyses (see below) supported the recognition of these taxa as separate species.
Molecular analyses. After alignment, the COI fragment had 618 nucleotide sites, of which 167 were parsi-mony–informative, and contained no insertions or deletions. A maximum likelihood tree extracted from 10 species of the subgenus Panonychus s.str., including P. caricae , plus two species of the subgenus Sasanychus in the genus Panonychus based on the COI gene of mtDNA indicated that these 12 species are clearly separated from each other with high bootstrap values (75–100%) ( Fig. 22 View FIGURE 22 ). Panonychus osmanthi morphologically resembles P. citri but produced sterile F 1 females when mated with P. citri ( Kitashima & Gotoh 1995; Ehara & Gotoh 1996; Ehara 1999). These two species formed a cluster with high bootstrap value (100%), the same as described in the previous molecular phylogenetic analyses ( Toda et al. 2000; Matsuda et al. 2014, 2018). The re-described species P. caricae formed a monophyletic clade with P. ulmi with a 95% bootstrap value. Panonychus hadzhibejliae was located at the root of the same clade with a 92% bootstrap value. Based on a morphological study, Arabuli et al. (2016) reported that P. hadzhibejliae is a valid species, and not a synonym of P. citri , and this is confirmed here with molecular data.
A comparison of the genetic distances of the COI gene showed that intra-specific divergence of the two P. mori strains (voucher specimen nos. 239 and 663) was higher (4.05%) than inter-specific divergence among P. citri — P. osmanthi (0.97%) and P. caricae — P. ulmi (1.94 and 2.10%) ( Table 2 View TABLE 2 ). However, these two strains of P. mori formed the same cluster with high bootstrap value (100%) and clearly separated from other species.
In the COI tree ( Fig. 22 View FIGURE 22 ), the two subgenera, Panonychus s.str. and Sasanychus , comprise strongly supported monophyletic clades with 98% bootstrap values. Sasanychus is treated as a subgenus of Panonychus by Bolland et al. (1998) and in the ‘Spider Mites Web’ database ( Migeon & Dorkeld 2006 ‾2020). This result supports the genusstatus given to Sasanychus by Ehara & Gotoh (1987), a result further supported by previous molecular phylogenetic studies ( Matsuda et al. 2014, 2018) and morphology. Sasanychus differs from Panonychus by the dorsal idiosomal setae not set on tubercles, the opisthosoma with transverse striae in the dorsocentral area, and nine and eight tactile setae on tibiae I and II, respectively (see Introduction for details). Thus, the genus Sasanychus is resurrected based on molecular and morphological evidence.
......continued on the next page

| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |