
Characidium summus, Zanata, Angela M. & Ohara, Willian M., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4021.2.7 |
|
publication LSID |
lsid:zoobank.org:pub:76B316C9-F7F8-4659-8693-B9771F34FFD2 |
|
DOI |
https://doi.org/10.5281/zenodo.6102840 |
|
persistent identifier |
https://treatment.plazi.org/id/03C58785-FF8B-1720-78EF-3A96FEF3F99A |
|
treatment provided by |
Plazi |
|
scientific name |
Characidium summus |
| status |
sp. nov. |
Characidium summus View in CoL , new species
( Fig. 1 View FIGURE 1 a)
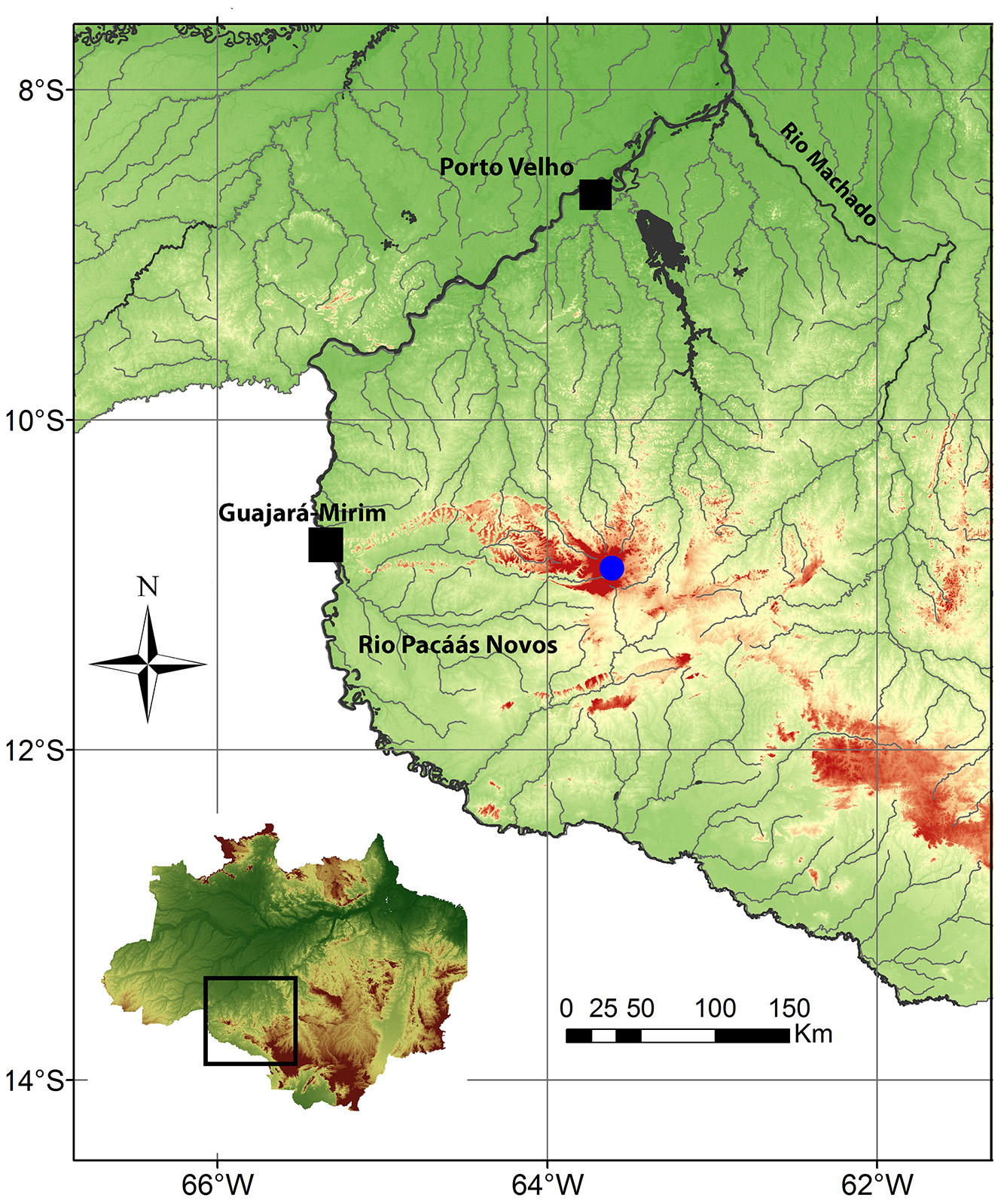
Holotype. MZUSP 116954, 34.5 mm SL, Brazil, Rondônia, Guajará-Mirim, upper rio Pacaás Novos, tributary of rio Mamoré, rio Madeira basin, 10º50'46.29"S 63º37'47.29"W, D. Hungria & A.C. Ribeiro, 17 Dec 2013.
Paratypes. MZUSP 116105, 11, 1 CS, 14.0– 42.3 mm SL; MNRJ 42909, 3, 25.6–39.4 mm SL; INPA 46986, 3, 28.2–30.9 mm SL; UFRO 23255, 2, 14.3–29.2 mm SL, collected with holotype.
Diagnosis. Characidium summus is a unique species among congeners by lacking an adipose fin and having a complete lateral line. It can be further distinguished from congeners, including C. vestigipinne Buckup & Hahn , that have complete lateral line and adipose fin absent, vestigial or reduced by the absence of dark bars or spots on head, body or fins, except for a dark blotch immediately on the rear of opercle, a narrow midlateral dark stripe (less than half scale deep), and a dark basicaudal spot.
Description. Morphometric data of holotype and paratypes presented in Table 1. Body fusiform and moderately compressed. Greatest body depth at vertical through dorsal-fin origin. Dorsal profile convex from upper lip to vertical through nares, slightly convex to nearly straight from this point to end of occipital process, slightly convex to nearly straight from this point to origin of dorsal-fin base, slightly convex along dorsal-fin base, and nearly straight from end of dorsal-fin base to origin of anteriormost dorsal procurrent caudal-fin ray. Vent ral profile straight to slightly convex from anterior portion of head to pelvic-fin origin, straight from latter point to origin of anal fin, straight and posterodorsally slanted along anal-fin base and straight along caudal peduncle. Snout triangular-shaped in lateral view. Mouth subterminal, slightly more dorsal than ventral edge of orbit. Distal tip of maxilla surpassing anterior margin of orbit. Orbit rounded, horizontal length longer than snout length. Cheek thin, its depth around a third to a quarter of orbit diameter. Nares separated; anterior naris with raised margins; posterior naris considerably closer to orbit than to anterior naris, without skin flaps. Supraorbital reduced to a minute ossification on anterodorsal portion of eye ( Fig. 2 View FIGURE 2 ). Nasal bones restricted to the ossified canal, without lateral lamella. Parietal fontanel limited anteriorly by frontals. Parietal branch of supraorbital canal present, not reaching frontal-parietal border. Epiphyseal branch of supraorbital canal absent.
Dentary teeth in two rows; outer series with 10(1) or 12(1) conical teeth; teeth decreasing in size from symphysis; inner series, with around 15 minute conical teeth inserted on edge of replacement tooth trench. Dentary teeth usually inclined anteriomedially; distal portion of symphyseal tooth overlapping its contralateral in some specimens. Premaxilla with a single series of 7(2), 8*(12), 9(2), or 10(1) conical teeth, decreasing in size from symphysis. Maxillary edentulous. Ectopterygoid teeth 6(1), conical, distributed in one series. Mesopterygoid teeth absent.
Scales cycloid; circulii absent and around 25 radii present on posterior field of scales. Lateral line complete, perforated scales 35(3), 36*(11), or 37(3); total horizontal scale rows above lateral line 4*(15) or 5(2); horizontal scale rows below lateral line 5*(14) or 6(2). Scales along middorsal line between supraoccipital and origin of dorsal fin around 15, but mostly disordered. Scales rows around caudal peduncle 14*(17). Axillary scale absent. Isthmus completely scaleless, but area between pectoral fins covered by scales. Pseudotympanum present, represented by small muscular hiatus at vertical through anterior portion of the swimbladder; hiatus situated between ribs of fifth and sixth vertebrae ( Fig. 3 View FIGURE 3 ).
Dorsal-fin rays ii,9(3), 3,i,8(1), or 3,i,9*(13); distal margin of dorsal fin rounded. Dorsal-fin origin at middle of standard length and slightly anterior to vertical through pelvic-fin origin. Adipose fin absent (20). Pectoral-fin rays highly variable iii,6,i(2), iii,6,ii(2), iii,7,i(2), iii,8,i(1), iii,8,ii(1), iv,6,i(1), iv,6,i(2), iv,7(4), or 4,i,8*(1); 1st and 2nd branched pectoral-fin rays longest; posterior tip of pectoral fin not reaching pelvic-fin insertion. Pelvic-fin rays i,6,i*(16), or i,7(1); 2nd and 3rd branched pelvic-fin rays longest; posterior tip of pelvic fin not reaching anal-fin origin. Anal-fin rays ii,6*(15), iii,6(1), and iv, 5 in the CS specimen; posterior margin of anal fin rounded. Caudalfin rays i,7,8,i(1), i,8,8,i*(15), or i,9,9,i(1). Caudal fin forked, lobes somewhat pointed, of similar size. Dorsal procurrent caudal-fin rays 9(1); ventral procurrent caudal-fin rays 9(1).
Total number of vertebrae 36(1); precaudal vertebrae 21(1); caudal vertebrae 15(1). Supraneural bones 4(1), similar in size. Epural bones 3(1). Uroneural bones 1(1). Branchiostegal rays 4(1); 3 connected to anterior ceratohyal, 1 connected to area between anterior and posterior ceratohyal.
TABLE I. Morphometric data of holotype and paratypes of Characidium summus (number of specimens measured = 17). The range includes the holotype. SD = standard deviation.
Color in alcohol. Ground color of head and body light brown ( Fig. 1 View FIGURE 1 a). Head with minute melanophores homogeneously distributed over dorsal and ventral portion, not forming spots. Body similarly colored, usually slightly darker on dorsal half. Some specimens with inconspicuous reticulate pattern on dorsal half of body due to light concentration of melanophores on posterior margin of scales. Dark midlateral narrow stripe (less than half scale deep) usually extending from rear of opercle to caudal peduncle, but not reaching its end or the basicaudal spot. Dark blotch immediately at rear of opercle, over two first scales of the lateral line and formed by pigment on skin; blotch somewhat round in most specimens. Smallest specimen examined without dark stripe and anterior blotch. Vertical dark bars on body absent (n=19); one specimen with very inconspicuous vertical blotches below dark midlateral stripe on first half of body. Basicaudal black spot conspicuous and well defined, somewhat vertically elongated, occupying area on the base of 4–6 caudal-fin rays; small specimens with basicaudal spot distinctly black. Fins similarly colored to each other, with melanophores over rays; interradial membranes hyaline; paired and anal fins somewhat less pigmented on its distal half.
Etymology. From the Latin summus , which means high, elevated, alluding to the high altitude where the species occurs, near Pico Tracoá, higher than 1100 m above sea level in the Serra dos Pacaás Novos.
Sexual dimorphism. No hooks on fins or other sexually dimorphic features were observed externally on the specimens examined.
Distribution. The new species is only known from upper rio Pacaás Novos, a tributary of rio Mamoré, rio Madeira basin, Rondônia State, Brazil ( Fig. 4 View FIGURE 4 ).
Habitat and ecological notes. Characidium summus occurs in a headwater of the rio Pacaás Novos, at 920 meters above sea level, situated on the Serra dos Pacaás Novos, close to the highest peak of the Rondônia State, named Pico Tracoá, higher than 1100 m. The type locality of C. summus is within the Conservation Unit Parque Nacional de Pacaás Novos, situated in the central portion of Rondônia State. The rio Pacaás Novos is a black water river, with moderate water current and predominantly rocky bottom. Specimens of C. summus were sampled in small puddles (1–4 m wide and 1.5 m deep) and laterally in the river backwaters, with water temperature of 22ºC, pH 3.58, dissolved oxygen 7mg /L, satured oxygen 16%, and conductivity 10µm/cm. Only juveniles of Erythrinus erythrinus (Bloch & Schneider) were sampled syntopically with the new species. Both species have color pattern and general shape of the body remarkably similar ( Fig. 1 View FIGURE 1 b).
Remarks. Characidium summus does not fit into any group of Characidium previously proposed in the literature (e.g., Buckup & Reis, 1997; Buckup & Hahn, 2000; Graça & Pavanelli, 2008; Netto-Ferreira et al., 2013). The unique potential synapomorphy shared with clades within Characidium proposed by Buckup (1993b) is the scaleless isthmus, one of the features of the Clade C1. The absence of adipose fin is shared with C. mirim Netto-Ferreira, Birindelli & Buckup , C. nupelia da Graça, Pavanelli & Buckup, C. stigmosum Melo & Buckup , C. xavante da Graça, Pavanelli & Buckup, and C. vestigipinne . However, according to the hypothesis of Netto- Ferreira et al. (2013), these species belongs to the Clade C4, supported by two synapomorphies not observed in Characidium summus : absence of the parietal branch of the supraorbital canal and absence of the inner row of teeth on dentary. Furthermore, the new species does not share the incompleteness of lateral line or color patterns of C. mirim , C. nupelia , C. stigmosum , and C. xavante . The combination of adipose fin absent and lateral line complete occurs occasionally in specimens of C. vestigipinne . According to the original description, in C. vestigipinne the adipose fin can be absent, vestigial or reduced in size ( Buckup & Hahn, 2000). However, as stated above, the latter species is a member of the Clade C4, closely related to C. rachovii Regan , C. stigmosum , C. occidentale Buckup & Reis, and C. orientale Buckup & Reis , sharing presence of a dark band along distal margin of anal and pelvic fins in adult males ( Netto-Ferreira et al., 2013).
A noteworthy condition for a species of Characidium observed in C. summus is the almost complete absence of supraorbital ( Fig. 2 View FIGURE 2 ). In fact, a minute vestigial supraorbital occurs. Loss of supraorbital was used to defined suprageneric relationships within characidiins and proposed as one of the three synapomorphies supporting the sister-group relationship of Microcharacidium and Odontocharacidium , the hypothesis B of Buckup (1993b). The other two synapomorphies are loss of postcleithrum 1 and complete loss of lateral tooth cusps, the former not occurring in C. summus and the latter present. Although having a vestigial supraorbital, a feature not observed in congeners, C. summus possess the conspicuous basicaudal dark spot and overall body morphology similar to the species included in Characidium .
The new species possesses a remarkably small pseutotympanun when compared to the structure described recently for C. bahiense Almeida , C. bimaculatum Fowler , C. samurai Zanata & Camelier , and C. timbuiense Travassos ( Zanata & Camelier, 2014) and other species examined herein for this feature. In C. summus , the pseudotympanum is barely or not visible externally by transparency. After removal of the skin the aperture observed is elongate and narrow, positioned between ribs of the fifth and sixth vertebrae, which remain concealed ( Fig. 3 View FIGURE 3 ). The opposite condition, with pseudotympanum visible through body wall and represented by a distinctly large aperture, usually with portions of two ribs exposed, occurs in species of Characidium with a reduced lateral line (e.g., C. bahiense , see Zanata & Camelier, 2014: fig 3). This is the case in C. mirim and C. xavante , both with pseudotympana similar to C. bahiense but with an even larger aperture, most of the hiatus situated between ribs of the fifth and sixth vertebrae plus a small opening anterior to the rib of fifth vertebra or posterior to the rib of the sixth vertebra. Characidium nupelia and C. stigmosum also possess similar pseudotympana, although with aperture slightly smaller.
According to Bockmann & Castro (2010), a large and widely exposed pseudotympanum seems to be characteristic of small-sized otophysan fishes, which inhabit calm waters, including a few species of Characidium . Comparisons of aspects of the pseudotympanum and lateral line completeness in species of Characidium inhabiting lotic and lentic environments reveal possibly related characters. As discussed above, examined species of Characidium with lateral line incomplete also possess a widely exposed pseudotympanum and inhabit relatively calm waters (e.g., C. bahiense , C. mirim , C. nupelia , C. xavante , and C. stigmosum ), and congeners inhabiting moderate to fast running waters possess a complete lateral line and comparatively smaller pseudotympanum (e.g., C. bimaculatum , C. samurai , C. summus , and C. timbuiense ). However, clear-cut criteria to indicate the degree of dependence between these features are lacking; confirmation of the hypothesis presented herein depends on future studies.
Characidium summus View in CoL apparently is a moderate water current dwelling fish and does not possess adaptations cited by Buckup et al. (2000) for congeners that inhabit fast water current environments, as a streamlined body and paired-fins modifications. Rather, somewhat fragile paired fins and a somewhat truncated body are observed in the new species. As informed elsewhere, only juveniles of Erythrinus erythrinus were sampled syntopically with C. summus View in CoL . Both species have a color pattern and general body shape remarkably similar ( Fig. 1 View FIGURE 1 a, b), interpreted herein as a putative mimetic association among them. Mimetic relations involving Characidium View in CoL , particularly C. heirmostigmata View in CoL , was previously proposed by Tencatt et al. (2014) in the description of the syntopic catfish Corydoras lacrimostigmata Tencatt, Britto & Pavanelli, 2014 View in CoL . According to the authors, both species possess resemblance in color pattern and form of caudal-fin lobes, more rounded in C. lacrimostigmata View in CoL than in congeners and very similar to the caudal-fin lobes of C. heirmostigmata View in CoL . Erythrinus erythrinus possess widespread distribution, predatory habits, and a somewhat similar coloration pattern throughout South America ( Oyakawa et al., 2013), possibly representing a model of Batesian mimicry. Characidium summus View in CoL , on the other hand, is apparently narrowly distributed, possess distinct color pattern from congeners, and possibly represents the mimic in the association. Previous mimetic relations involving E. erythrinus were described by Brosset (1997). However, that is a case of aggressive mimicry where E. erythrinus plays the mime and Laimosemion agilae (Hoedeman) View in CoL the model, in a male-oriented mimicry. Understanding the ecological relationships between C. summus View in CoL and E. erythrinus is critical to assess its ecological significance and the possible evolutionary process involved in the association.
Comparative material examined. All from Brazil, except when noted. Characidium alipioi : MNRJ 5550, holotype, 50.0 mm SL, Rio de Janeiro, rio Paraíba do Sul basin; MNRJ 5551, paratype, 51.0 mm SL; MNRJ 5552, paratype, 48.3 mm SL; MNRJ 5553, paratype, 45.0 mm SL. MZUSP 112331, 6, 50.3–72.3 mm SL, São Paulo, rio Paraíba do Sul basin. MZUSP 80224, 12, 36.5–61.9 mm SL, Rio de Janeiro, rio São João basin. Characidium bahiense : MZUSP 8940, holotype, 16.0 mm SL, Bahia, Arembepe; MZUSP 8923, paratype, 18.7 mm SL; MZUSP 8924, paratype, 21.5 mm SL. UFBA 7167, 23, 3 CS, 21.1–26.0 mm SL, Bahia, rio Itapicuru basin. Characidium bimaculatum : MNRJ 21249, 14, 20.5–41.4 mm SL, Ceará, rio Curu basin. MNRJ 4925, 1, 25.5 mm SL; MNRJ 4928, 1, 28.7 mm SL, Ceará, rio Salgado basin. MZUSP 110779, 8, 29.4– 24.3 mm SL, Paraíba, rio Acaraú basin.
UFBA 3829, 6, 1 CS, 22.5–30.8 mm SL, Paraíba, rio Piranhas basin. MZUSP 68959, 1 CS, 22.2 mm SL, Rio Grande do Norte, rio Ceará-Mirim. Characidium cf. declivirostre : MZUSP 97137, 1, 35.4 mm SL, Pará, rio Xingu basin. MZUSP 82084, 2, 44.5–46.5 mm SL, Mato Grosso, rio Juruena basin. MZUSP 84965, 1CS, 43.8 mm SL, Amazonas, rio Negro basin. Characidium etheostoma : MZUSP 96860, 2CS, 40.7–47.1 mm SL, Pará, rio Xingu basin. Characidium fasciatum : MZUSP 39676, 15, 32.7–36.4 mm SL, Minas Gerais, rio São Francisco basin. Characidium gomesi : MZUSP 73193, 47, 25.8–32.0 mm SL, Minas Gerais, rio Paranaíba basin. MZUSP 88440, 3, 29.5–42.8 mm SL, São Paulo, rio Corumbataí. MZUSP 98815, 1CS, 52.4 mm SL, São Paulo, rio Tietê. Characidium grajahuensis : MNRJ 3855, holotype (snout damaged, precise measurement currently not possible), Rio de Janeiro, Grajaú. Characidium hasemani : MZUSP 91785, 4, 44.0–58.0 mm SL, Mato Grosso, rio Xingu basin. Characidium heirmostigmata : MZUSP 97738, holotype, 34.6 mm SL, Paraná, rio Paraná basin. Characidium interruptum : MZUSP 58992, 1, 30.9 mm SL, Rio de Janeiro, rio São João basin. Characidium japuhybense : MNRJ 5194, holotype (snout damaged, precise measurement currently not possible), Rio de Janeiro, Angra dos Reis. Characidium lagosantense : MNRJ 3852, holotype (snout damaged, precise measurement currently not possible), Minas Gerais, rio São Francisco basin. MNRJ 18108, 71, 11.2–31.2 mm SL, Minas Gerais, rio São Francisco basin. Characidium lanei : MNRJ 6185, holotype, 40.8 mm SL, São Paulo, rio Ribeira do Iguape basin; MNRJ 9766, paratype, 36.2 mm SL. MNRJ 32884, 38, 20.9–36.4 mm SL, São Paulo, rio Lajeado. MZUSP 69585, 7, 25.2–31.0 mm SL, São Paulo, rio Ribeira do Iguape basin. Characidium laterale : MZUSP 90204, 2, 20.6–25.1 mm SL, Mato Grosso, rio Paraguai basin. MZUSP 96687, 80, 15.7–24.0 mm SL, Mato Grosso, rio Paraguai basin. Characidium lauroi : MNRJ 5529, holotype, 60.4 mm SL, Rio de Janeiro, rio Paraíba do Sul basin; MNRJ 5530, paratype, 56.8 mm SL. MZUSP 110359, 5, 31.1–45.9 mm SL, São Paulo, rio Paraíba do Sul basin. Characidium mirim : MZUSP 97724, paratypes, 4, 18.9–19.7 mm SL, Mato Grosso, rio Araguaia basin. Characidium nupelia : MZUSP 87743, holotype, 29.0 mm SL, Mato Grosso, rio Paraguai basin. Characidium oiticicai : MNRJ 9480, holotype, 35.3 mm SL, São Paulo, rio Tietê basin. MZUSP 108640, 2, 37.4–38.1 mm SL, São Paulo, rio Tietê basin. MZUSP 26973, 5CS, 37.4–51.3 mm SL, São Paulo, rio Tietê basin. Characidium cf. pteroides : MZUSP 85653, 81, 16.3–28.7 mm SL, Amazonas, rio Negro basin. Characidium pterostictum : MZUSP 43547, 15, 28.7–37.8 mm SL, Rio Grande do Sul, Mampituba system. Characidium rachovii : MZUSP 49163, 8, 25.9–34.2 mm SL, Rio Grande do Sul, lagoa dos Patos drainage. MZUSP 49172, 1CS, 26.9 mm SL, Rio Grande do Sul, rio Grande. Characidium samurai : MZUSP 108188, holotype, 46.6 mm SL, Bahia, rio das Almas basin; MZUSP 112385, paratypes, 9, 1 CS, 25.4–42.7 mm SL; UFBA 7259, paratypes, 7, 20.8–42.6 mm SL, collected with holotype. Characidium stigmosum : MZUSP 40804, holotype, 33.5 mm SL, Goiás, rio Tocantins basin. Characidium tenue : MZUSP 63803, 6, 31.0– 42.2 mm SL, Rio Grande do Sul, rio Jacuí basin. Characidium timbuiense : MNRJ 4285, holotype, 51.8 mm SL, Espírito Santo, rio Timbuí basin. Characidium vidali : MNRJ 9757, holotype, 47.8 mm SL, Rio de Janeiro, rio Soberbo. MZUSP 893909, 2 CS, 46.9–48.4 mm SL, Rio de Janeiro, rio São João. Characidium xanthopterum : MZUSP 53404, 2CS, 35.0– 35.9 mm SL, Goiás, rio Tocantins. Characidium xavante : MZUSP 87745, paratypes, 20, 13.9–22.7 mm SL, Mato Grosso, rio Xingu basin. MZUSP 91718, 2CS, 118.6– 19.1 mm SL, Mato Grosso, rio Xingu basin. Characidium cf. zebra : MZUSP 92910, 7, 26.5– 29.0 mm SL, Amazonas, rio Negro basin. MZUSP 98203, 11, 26.5–31.4 mm SL, Mato Grosso, rio Xingu basin. MZUSP 85091, 2, 52.0– 57.8 mm SL, Amazonas, rio Negro basin. Peru. Characidium cf. zebra : MZUSP 77838, 2, 23.5–27.1 mm SL, Loreto, rio Pastaza basin. Guiana. Characidium amaila : MZUSP 109096, 1CS, 64.9 mm SL, Potaro-Siparuni, rio Kuribrong. Venezuela. Characidium cf. zebra : MZUSP 96477, 1, 35.0 mm SL, Bolivar, rio Orinoco basin.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Characidium summus
| Zanata, Angela M. & Ohara, Willian M. 2015 |
Corydoras lacrimostigmata
| Tencatt, Britto & Pavanelli 2014 |