
Neacomys Thomas, 1900
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4920.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:F898059E-0B23-48F1-BFAE-BBBB3D87DA5A |
|
DOI |
https://doi.org/10.5281/zenodo.4531858 |
|
persistent identifier |
https://treatment.plazi.org/id/03C68783-7D67-1038-FF29-7E3A8868FBF0 |
|
treatment provided by |
Plazi |
|
scientific name |
Neacomys Thomas, 1900 |
| status |
|
Genus Neacomys Thomas, 1900
Neacomys serranensis , new species ( Figures 1-12 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 ; Tables 1-5)
Serrano Spiny Mouse or Ratoncito Espinoso Serrano
Holotype. — A young adult male specimen (age class II) housed in the Colección de Mamíferos del Museo de Historia Natural de la Universidad Industrial de Santander (UIS-MHN-M 1608), caught in an entomological pitfall trap by Alejandro Lopera and prepared by Javier E. Colmenares-Pinzón in February 28, 2018 (original field number JEC 127 ); skin with skull and mandibles, in good condition; liver, heart, and skeletal muscle fragments preserved in 96% ethanol and maintained in the Colección de Tejidos del Museo de Historia Natural UIS; baculum preserved in 10% buffered formalin; ectoparasites preserved in 70% ethanol.
Type locality. — Colombia, Department of Santander, Municipality of El Carmen de Chucurí , vereda La Bodega, sector Manchurrias, western slope of the Serranía de los Yariguíes, Finca Buenos Aires, in the basin of the La San Guillerma stream (6°41’5.2’’N, 73°26’10.4’’W, 1,595 m a.s.l.) ( Figs. 1 View FIGURE 1 and 5A View FIGURE 5 ) GoogleMaps .
Referred specimens. — Only an additional very young specimen, also caught in an entomological pitfall trap by Juliette Gualdrón in November 25, 2019, and preserved in fluid (ethanol 96%) with not extracted skull (UIS-MHN-M 1928); tissues (fragments of heart and liver) preserved in ethanol 96%. This specimen was collected southern of the type locality (̴13.45 Km), in the eastern slope of the Serranía de los Yariguíes [ Colombia, Department of Santander, Municipality of Hato, vereda Hoya Negra (6°35’37.81’’N, 73°21’19.65’’W, 2,124 m a.s.l.)] ( Figs. 1 View FIGURE 1 and 5B View FIGURE 5 ) GoogleMaps .
Distribution and natural history. — Neacomys serranensis sp. nov. is only known from two localities in opposite slopes of the Serranía de los Yariguíes, but its distributional limits are not well characterized yet. The Serranía de los Yariguíes is a massif located within the limits of the Department of Santander, in Northeastern Colombia, of about 80 Km South to North, emerging from the lowlands of the Magdalena Valley with an approximate area of 5,000 Km 2 and altitudes between 500 and 3,500 m ( Díaz, 2008; Jara-Muñoz & Zabala-Rivera, 2018) ( Figs. 1 View FIGURE 1 and 5C View FIGURE 5 ). It is considered as part of the Cordillera Oriental, the eastern ramus of the Colombian Andes, but is isolated from the main mountain chain by the dry valleys of the Suárez River in the east, and the Sogamoso River in the north. The entire massif belongs to the Magdalena Valley Montane Forest ecoregion ( Olson et al., 2001). The type locality of the species is at 1,595 m on the western slope, the less intervened portion (Donegan et al., 2009); it is dominated by terra firme high dense forest, and corresponds with the lower montane moist forest life zone and the Andean orobiome ( Holdridge, 2000; Diaz, 2008) ( Fig. 5A View FIGURE 5 ). The climate is characterized by a bimodal rainfall regime with a first peak of rains in April-May, and a second in September-October; December-February, as well as July, are the months with the lowest precipitation values. Mean annual precipitation is 1,625 mm and mean annual temperature is 18.89 °C ( Fick & Himans, 2017). Plant families with highest importance value in the zone are Burseraceae , Lauraceae , Clusiaceae , Moraceae , and Meliaceae (Ayala, 2011) . The holotype was caught on the left bank of the La San Guillerma stream, in an entomological pitfall trap baited with approximately 50 g of human feces and installed in a forest belt with abundant leaf litter and a canopy above 20 m. There, historical logging is evident, and the surrounding area has been transformed into crops (coffee) and pastures for livestock. The collected individual is a young adult male with no scrotal testis. Other small terrestrial mammals also were recorded in nearby forest, some of them comprising typical montane taxa: Cryptotis cf. tamensis, Handleyomys alfaroi, Heteromys australis, Marmosa (Micoureus) phaea, Marmosops caucae, Nephelomys childi, Oligoryzomys sp., Rhipidomys fulviventer , and Thomasomys laniger .
The second locality is at 2,124 m on the eastern slope of the massif, the most intervened portion. The landscape is dominated by a mosaic of pastures with scarce fragments of secondary growth forest and corresponds with the lower montane moist forest life zone and the upper Andean orobiome ( Holdridge, 2000; Diaz, 2008) ( Fig. 5B View FIGURE 5 ). The climate also is characterized by a bimodal regime with the first peak of rains between April and July, and the second in September-October. November-March are the months with the lowest precipitation values, as well as August. The mean annual precipitation is 1,503 mm, whereas the mean annual temperature is 16.08 C° ( Fick & Himans, 2017). The young specimen collected there was caught in a forest remnant along a small stream, in a pitfall trap baited with human feces. The surrounding area has been severely deforested and transformed into pastures for livestock that remain abandoned, which is evidenced by the presence of abundant ferns (genus Pteridyum).
Etymology. — the specific epithet harbors a double meaning. Firstly, it derives from the Spanish adjective serrano, from the sierra o serranía, as the species have only been recorded within the range of the Serranía de los Yariguíes, an important isolated massif that is part of the western slopes of the Cordillera Oriental of Colombia. Secondly, it is a tribute to Victor Hugo Serrano Cardozo, Professor of the Universidad Industrial de Santander, who has supported for more than 20 years the study of the biodiversity in the Department of Santander, Colombia. Without his motivation, the discovering of this new species had not been possible.
Diagnosis. — Neacomys serranensis sp. nov is a small species within the genus Neacomys distinguished from other similar-sized congeners, especially from N. tenuipes (the only other Trans-Andean species in Colombia) by the following combination of characters: ventral fur superficially ochraceous-buff, with individual gray-based hairs from chin to anus (the same pattern in the inner surface of the arms and legs), sides of the muzzle (above the mouth) with distinctive broad patches of ochraceous-orange hairs, tail much longer than the combined length of head and body, with small caudal scales bearing long hairs (under magnification and at least at the terminal half), short rostrum, nasals short and expanded anteriorly, narrow zygomatic plate, very shallow zygomatic notches, long incisive foramina (in relation to diastema length), almost reaching the anterior alveolus of M1, maxillary with small perforations at the posterior border of the incisive foramina, dorsal margin of the tegmen tympani exposed above the ectotympanic dorsal margin (leaving to a smaller postglenoid foramen), dorsal margin of the ectotympanic not reaching the mastoid tubercle (leaving to an open ectotympanic ring), short and broad hamullar process of the squamosal, short subsquamosal foramen, wide oval orbicular apophysis with its longitudinal axis forming an almost right angle with the manubrium of malleus, paraoccipital process slightly bifurcated and situated well above the ventral margin of the occipital condyle, M1 with narrow and divided anterocone, principal labial cusps (paracone, metacone) smaller than lingual cusps (protocone, hypocone), mesoflexus divided into a short labial cleft and a broad triangular fossette that forms a prominent landmark in the center of the tooth, broad and distally bifurcated metaflexus, round and laterally positioned mental foramen, and broad coronoid process.
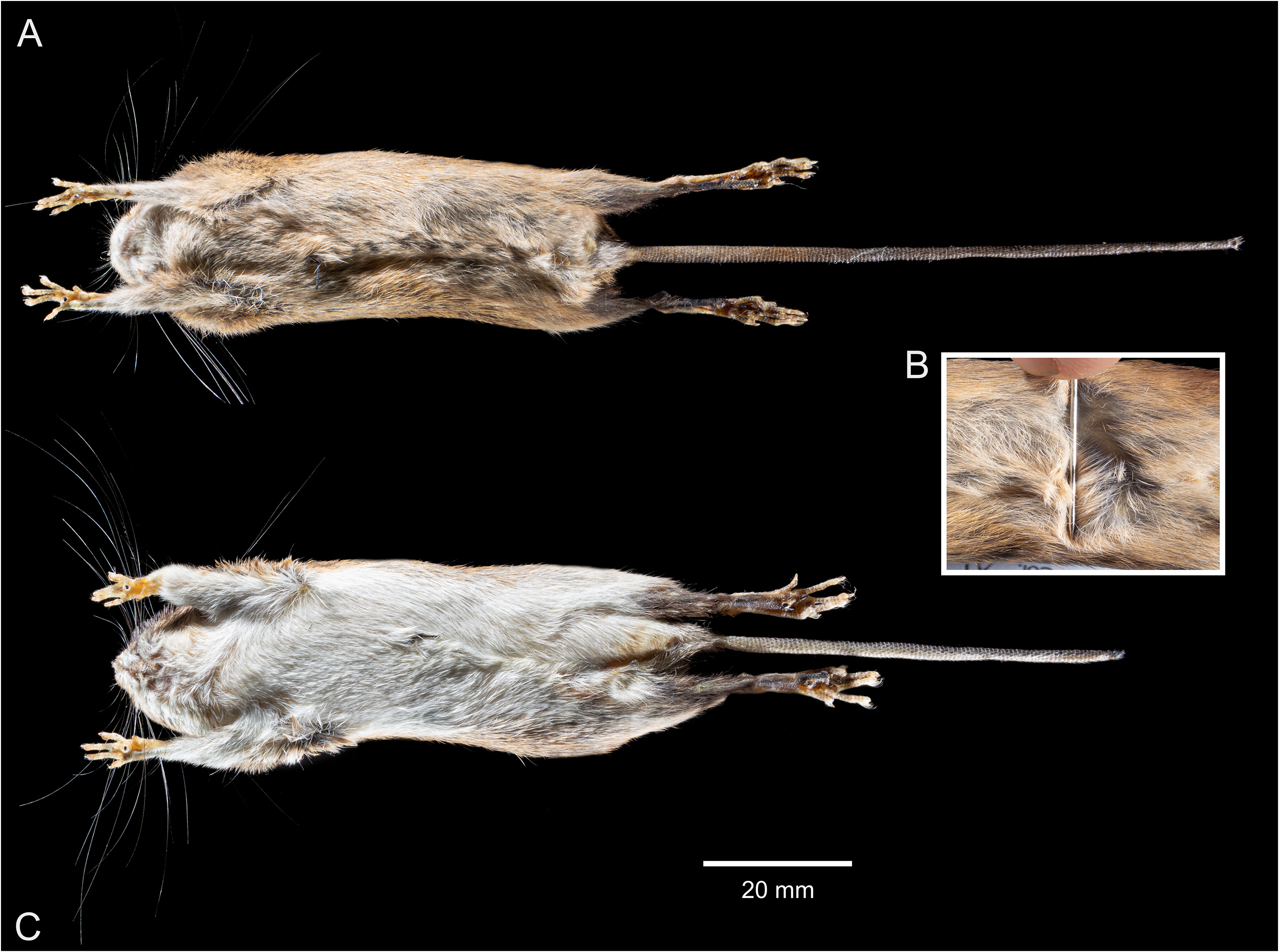
Morphological description — A small sized species. The dorsal pelage has a general orange brownish appearance given by a mix of individual predominantly orange hairs (with a very narrow blackish tip), and black-tipped spines ( Figs. 6A View FIGURE 6 and 12 View FIGURE 12 ). The brownish tint prevails in the middle of the back and the rump due to the concentration of black-tipped spines, while the paler orange tint prevails along the sides; most of the dorsal hairs and spines are grayish basally. The upper surface of the arms is covered by brown-grayish short hairs (some of them with a narrow buff-ochraceous tip) intermixed with buff-ochraceous hairs, which gives a general grayish appearance; the upper surface of the legs is similar to the adjacent parts of the dorsum. The ears are externally dark brown and scantily covered by dark-brownish and yellowish-orange short hairs; the region surrounding the external auditory meatus has a yellowish tint. The muzzle appears darker above (even more above the nose) due to the presence of entirely short black hairs. There are relatively broad orange-ochraceous patches in the sides of the muzzle (above the mouth) covering the area where the thickest mystacial vibrissae arise ( Fig. 7A View FIGURE 7 ). A clear narrow band, also ochraceousorange in color, delineates dorsal and ventral pelage. Ventral fur is superficially ochraceous-buff, with individual hairs grayish basally; the chin, the inguinal region, and the inner surface of the arms display a slightly paler tone ( Fig. 8A View FIGURE 8 ). The genal, superciliary, and mystacial vibrissae are long and extend beyond the posterior border of the pinnae. Dorsal surfaces of manus and pes are covered predominantly by short white hairs, but there are also white hairs with grayish-brown roots and entirely grayish brown hairs. Ungual manus tufts are whitish and do not pass the claws. Hind feet are small and narrow with six plantar pads; whitish ungual pedal tufts pass slightly the claws. Outer pedal digits are short, with the claw of the first digit reaching the second digit base, and the claw of the fifth digit extending approximately half length of the phalange 1 of adjacent fourth digit. Tail longer than the combined length of head and body (± 28% longer in the holotype), appearing dark brown dorsally but ventrally with a paler tone at the base. Relatively small caudal scales (20 per centimeter at the base of the holotype’s tail dorsal surface) are present, the dorsal ones bearing dark hairs and the ventral ones bearing paler (whitish) hairs. Tail upper hairs cover approximately the length of one scale at the first 1/3 of the total tail’s length, but in the remaining 2/3 they cover the length of one and a half scale; lower tail hairs are longer, covering more than the length of one scale (up to two scales) along the entire surface of the tail. There is an inconspicuous tuft of hairs at the tail’s tip.
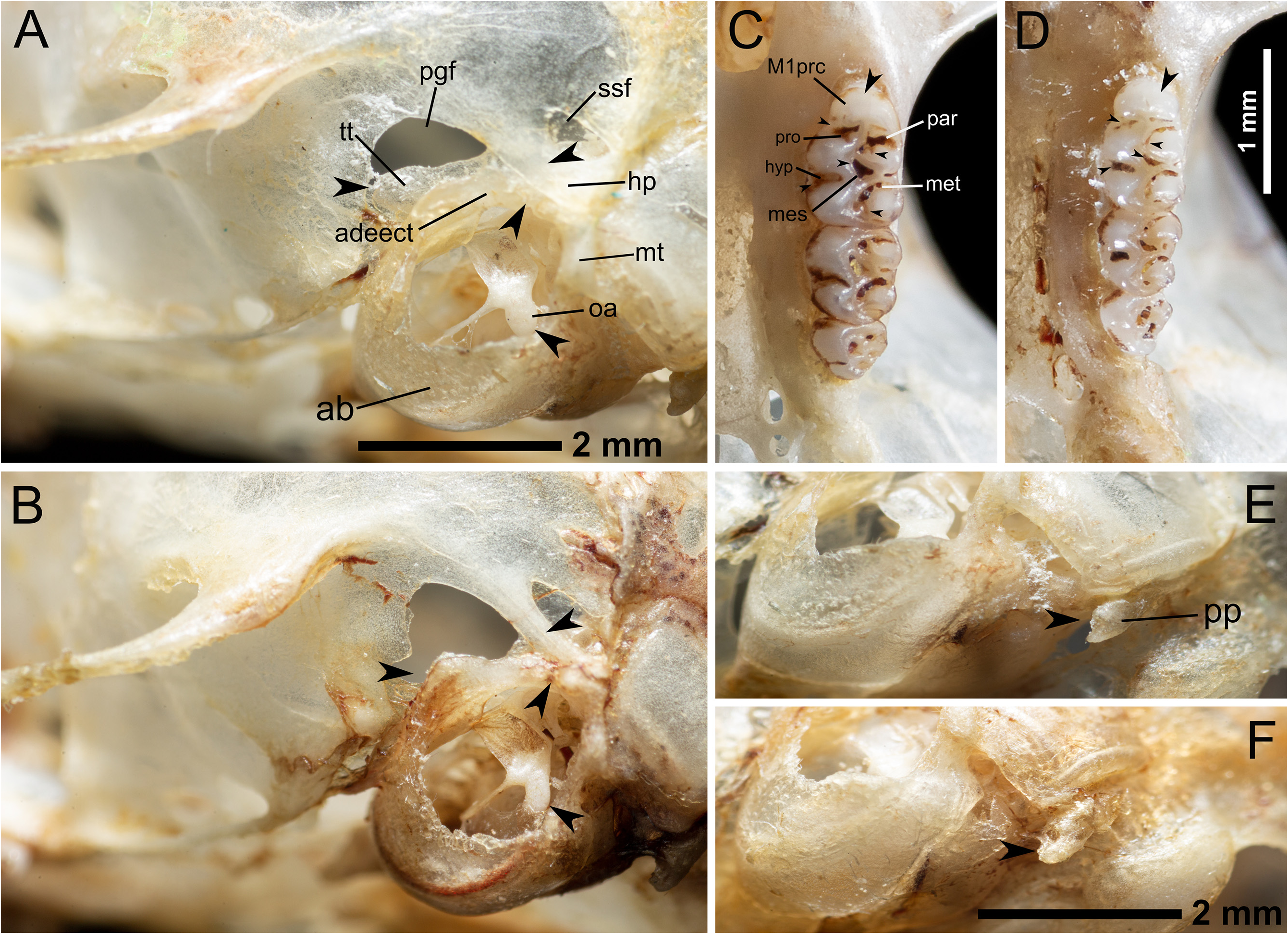
Skull with relatively short rostrum flanked by shallow zygomatic notches ( Fig. 9A View FIGURE 9 ). Short nasal bones expanded anteriorly, with irregular almost squared posterior ends placed slightly beyond the premaxillary-maxilary-frontal, and maxillary-frontal-lacrimal sutures; there is a slight depression at the end of the nasal bones. Sutures between maxillary and frontal bones appear as large interdigitations. Lacrimals are conspicuous and irregular in form, having a wide portion in contact with maxillary, and a small portion with a spinous projection in its posterior borders in contact with frontals. Interorbital region hourglass in shape, with supraorbital beads weakly developed, dorsally oriented, and continuing onto braincase as low temporal crest (reaching the fronto-parietal joint). Braincase relatively wide and conspicuously inflated
In lateral profile the skull seems convex due to the globose braincase ( Fig. 10A View FIGURE 10 ). The anterior edges of the nasals extend slightly beyond the rostral processes of premaxilla; gnathic process is short. Zygomatic plate is narrow and zygomatic notches indistinct; anterior border of plate relatively flat and placed slightly in front of anterior margin of superior maxillary root of zygoma; posterior margin of zygomatic plate situated anterior to the alveolus of M1. The infraorbital foramen is narrow (frontal view). The insertion of the superficial masseter is found at the inferior zygomatic root, slightly behind the anterior margin of the zygomatic plate. The sphenopalatine foramen is placed at the level of the metacone of M1. Alisphenoid strut is absent. The presence of a conspicuous sphenofrontal foramen and a squamosal-alisphenoid groove on each side, in conjunction with the presence of the stapedial foramen, indicate a primitive carotid circulation (pattern 1 of Voss, 1988; Carleton & Musser, 1989). The hamular process of the squamosal is short but relatively thick and stout ( Figs 10A View FIGURE 10 , and 11 View FIGURE 11 ). Tegmen tympani is partially exposed above the dorsal surface of the ectotympanic, acting as a membrane that partially covers the ventral portions of the subsquamosal and posglenoid foramina; tegmen tympani does not reach the posterior process of the squamosal ( Fig. 11 View FIGURE 11 ). Postglenoid foramen is relatively small and semicircular; the subsquamosal foramen is also small, shortened antero-posteriorly, and placed below the squamosal root of zygomatic arch. There is no contact between the anterodorsal edge of the ectotympanic and the mastoid tubercle, which leads to an opened ectotympanic ring ( Fig. 11 View FIGURE 11 ). The orbicular apophysis of the malleus is wide and elongate (oval in shape), with its longitudinal axis forming an almost right angle with the manubrium ( Fig. 11 View FIGURE 11 ). Mastoid with a conspicuous but small dorsolateral fenestra. The paraoccipital process is small, robust, slightly bifurcated and placed close to the auditory bulla and well above the ventral margin of the occipital condyle.
The incisive foramina are moderately long (averaging about 65% of diastema length), with outer borders slightly curved and posteriorly convergent; their posterior margins extend almost to the anterior alveolus of M1 ( Figs. 9B View FIGURE 9 and 11 View FIGURE 11 ). The premaxillary portion of the septum is slightly narrower than the maxillary portion. There are small perforations on the maxillary adjacent to the posterior margins of the incisive foramina. Relatively large and deep posterolateral palatal pits are placed posterior to M3, surrounding the anterior margin of the mesopterygoid fossa; at least in the holotype, there are two pits in one side and one in the other. Mesopterygoid fossa, which doesn´t reach the posterior end of M3, with wide M-shaped anterior margin (non-evident palatal spine). Irregular sphenopalatine vacuities are present around basisphenoid and presphenoid. The pterygoids are slightly convergent posteriorly; the parapterygoid fossae are shallow and triangular, with small irregularly distributed perforations but not evident fontanellae. Borders of the pterygoid plates are moderately curved. The posterior opening of the alisphenoid canal is oval, and its length is about M3 breadth. Auditory bullae small and flask-shaped, with the exposed flange of periotic extending to internal carotid canal; eustachian tube short and wide. Anterior bullae process is in contact with the posterior margin of the pterygoid plate. Anterior border of the foramen magnum is wide, with not conspicuous notch.
Mandible with a wide and short condyle ( Fig. 10A View FIGURE 10 ). Thin, short, and claw-shaped coronoid process situated well anterior to the condyle, forming a u-shaped sigmoid notch. Masseteric crest placed below the protoconid of m1; capsular process forming a subtle rounded elevation that is placed slightly beyond the coronoid process. Mental foramen relatively large and laterally positioned.
Upper incisors are opisthodont, with rounded tips and pigmented anterior face (yellowish orange). Molar rows are parallel; principal labial cusps (paracone, metacone) smaller than lingual cusps (protocone, hypocone) ( Fig. 11 View FIGURE 11 ). First maxillary molar with narrow anterocone (which gives the tooth a general subtriangular appearance), divided by a weakly developed anteromedian flexus (only evident in the right M1 of the holotype) ( Figs. 9B View FIGURE 9 and 11 View FIGURE 11 ); procingulum is anteriorly rounded. The anteroflexus does not reach the labial surface, and instead, is a relatively elongated internal fossette; the anteroloph is narrow, almost perpendicular to the longitudinal plane of the tooth and does not reach the labial surface. Paraflexus prominent, broad, long, and deep, divided distally by the intrusion of the protolophule. The mesoflexus is divided into a short labial cleft and a large internal fossette that forms a very prominent landmark in the center of the tooth. The mesoloph crosses the tooth diagonally and reaches the labial cingulum; it is relatively broad near the median mure, then gets compressed by the mesoflexus (labial cleft) but enlarges again when reaches the labial surface (at this zone, the mesoloph is almost perpendicular to the longitudinal axis of the tooth). The metaflexus is also prominent, wide, long, deep, and “bifurcated” distally. The posteroflexus is a short internal fossette; posteroloph is moderately narrow and reaches the labial surface. The protoflexus and hypoflexus are very distinct and broad. M2 is approximately squared in occlusal outline; the anteroloph, mesoloph, and posteroloph are almost parallel (perpendicular to the longitudinal axis of the tooth) and reach the labial surface. The paraflexus is narrow proximally (near the labial surface) but enlarges distally (in the middle of the tooth, near the anterior mure). The mesoflexus of M2 is divided and resembles that of M1, with a short labial cleft and a large internal fossette. The metaflexus is prominent and deep, while the posteroflexus is a short internal fossette. The protoflexus is indistinct, while the hypoflexus is clearly differentiated (deep and long). M3 is small, half the size of M2, and subtriangular; the anteroloph is short and straight. The paraflexus forms a continuous fold with a labial short element of the mesoflexus, surrounding the paracone; there is a distal (in the center of the tooth) element of the metaflexus forming a prominent internal fossete.
In the lower dentition, the procingulum of m1 is narrow, which gives the tooth a general subtriangular appearance. The anteromedian flexid is almost indistinct, however, the anterolingual and anterolabial conulids are evident. There is no trace of the anteroflexid, and the anterolophid is fused with the anteroconid. The metaflexid is prominent and is fused with the mesoflexid, forming a deep and broad groove that surrounds the metaconid. The mesolophid is poorly developed and fused with the entoconid; the entoflexid does not reach the labial surface, and instead, it is reduced to a small internal fossette. The posteroflexid is prominent, deep, and very broad; the posterolophid is well developed and reaches the labial surface. The hypoflexid is divided by the ectostylid into an anterior broad and long element, and a posterior small cleft, both reaching the labial surface; the protoflexid also reaches the lingual surface and is remarkably broad. The anterolabial cingulum of m2 is well marked but slender, followed by a narrow, shallow, and not very long protoflexid. The mesoflexid is very broad, long, and slightly bifurcated on its anterior end; the mesolophid and the entoflexid are indistinguishable, fused with the entoconid. Contrary, the posteroflexid is prominent, deep and very broad; the posterolophid is evident and reaches the labial surface. The hypoflexid forms a prominent, deep and long basin on the tooth surface. On m3 the anterolabial cingulum is very narrow, and the protoflexid is indistinguishable. The protoconid, hypoconid, and metaconid are well defined, while the entoconid, as well as the mesolophid, are not evident. The mesoflexid is a semicircular, very prominent, and relatively deep fossette, contrary to the posteroflexid, which is a shallow, short and narrow element. The posterolophid is well defined. The hypoflexid is prominent, long, deep, and moderately broad.
Holotype measurements — HBL: 80; LT: 102; HF: 20; Ear: 15.70; CIL: 18.36; LD: 5.12; LM: 2.82; BM1: 0.91; LIF: 3.11; BIF: 1.57; BPB: 2.19; BZP: 1.65; LR: 6.3; LN: 7.56; LIB: 4.32; BB: 10.4; ZB: 10.95; Wt: 14 ( Table 1).
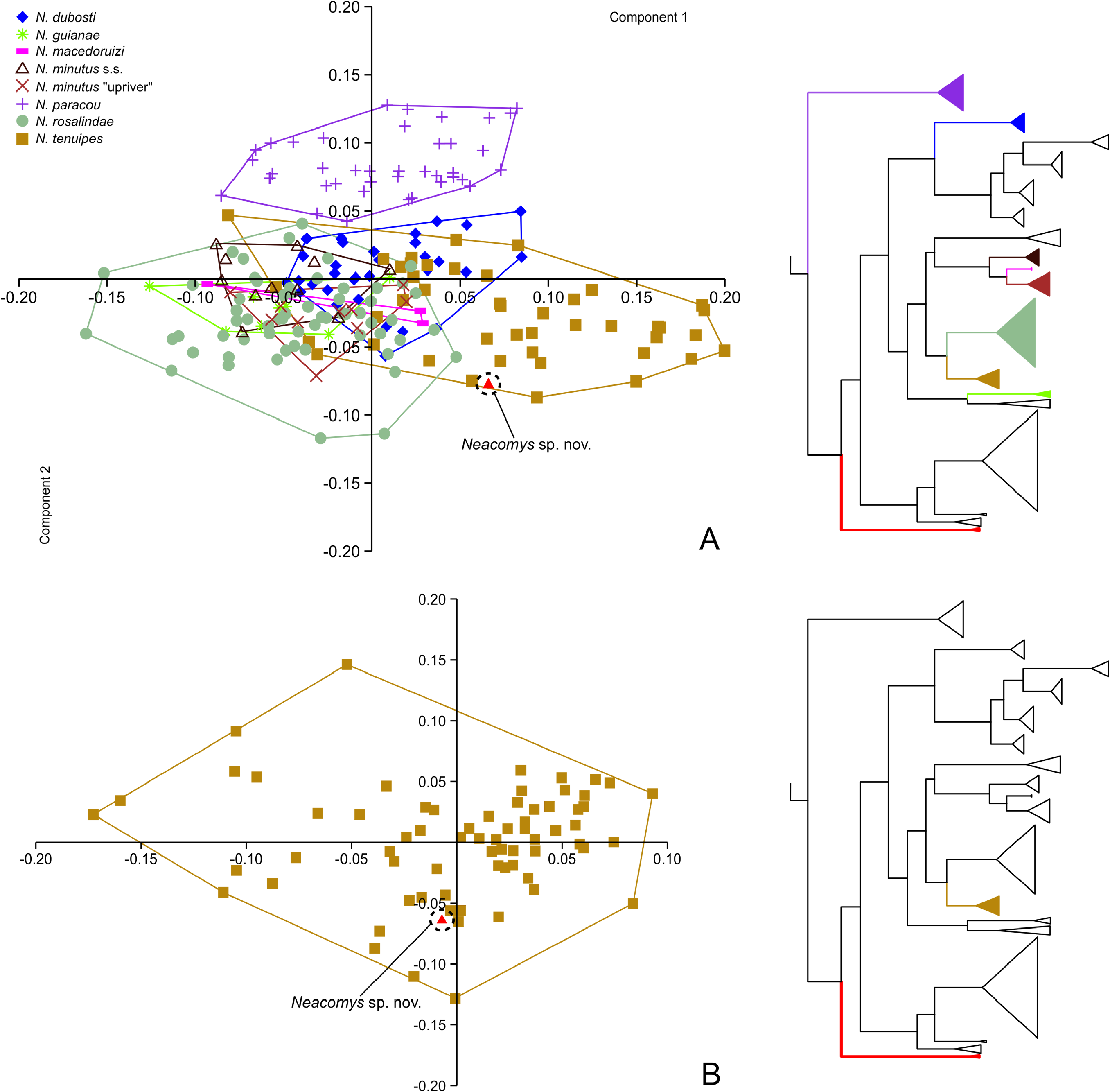
Comparisons — There are only two species of Neacomys confirmed for Colombia: a. The first of them, N. spinosus , have been reported from the departments of Meta and Vaupés, in the Amazonian lowlands east of the Cordillera Oriental (Cis-Andean) ( Solari et al., 2013). However, the taxonomical identity of these populations is uncertain according to the work of Hurtado & Pacheco (2017). These authors found remarkable genetical and morphological structure between different populations of N. spinous , recognizing at least three well discretized taxa: N. spinosus , N. vargasllosai , and N. amoenus . Relying on the distributional data provided in their work, but without the direct inspection of voucher specimens, it is possible to hypothesize that the Cis-Andean species of Neacomys present in Colombia is N. amoenus (subspecies N. a. carceleni), and not N. spinosus . Despite of the notably allopatric distribution between N. a. carceleni and N. serranensis sp. nov, is important to mention that both taxa are morphologically and morphometrically well differentiated; N. a. carceleni belongs to the monophyletic large-bodied group within the genus ( Sánchez-Vendizú et al., 2018) ( Fig. 3 View FIGURE 3 ) and its external and craniodental dimensions exceed those of N. serranensis sp. nov, which belongs to the non-monophyletic small bodied group (e.g. HF = 23.25 vs. 20; CIL = 21.78 vs. 18.36; LM = 3.20 vs. 2.82) ( Hurtado & Pacheco, 2017). Whereas N. serranensis sp. nov has a gray-based ochraceous-buff ventral fur, in N. a. carceleni the ventral coloration is completely white ( Table 3). The muzzle of N. serranensis sp. nov is also distinctive, with the presence of broad orange-ochraceous patches on each side (above the mouth) that have not been noted in specimens of N. a. carceleni ( Hurtado & Pacheco, 2017).
The lateral profile of the skull allows to discriminate between both species, being flat in N. a. carceleni and convex in N. serranensis sp. nov. due to its globose braincase; also, unlike N. a. carceleni, the tegmen tympani of the new species is not in contact with the posterior process of the squamosal. In ventral view, the most important differences between both species are probably at the level of the molar rows: these are parallelly oriented in N. serranensis sp. nov., with anteriorly narrowed first molars, whereas in N. a. carceleni they converge posteriorly and bear anteriorly rounded first molars. The mesoflexus of M1 and M2 are very distinctive in N. serranensis sp. nov., divided in a short labial cleft and a large internal fossette which are not present in upper molars of N. a. carceleni nor in any other species within the genus.
b. The other species of Neacomys distributed in Colombia is N. tenuipes ; according to Solari et al. (2013), Weksler & Bovicino (2015), and Sánchez-Vendizú et al. (2018) it has been reported west of the Cordillera Oriental in the departments of Antioquia, Boyacá, Caldas, Cauca, Chocó, Cundinamarca, Nariño, and Valle del Cauca, between 5 and 1,750 m. In the context of the present work, the species is also recorded from the departments of Bolívar (93 m; 105 m) and Santander (1,091 m) ( Fig. 2 View FIGURE 2 ; Appendix I). Although N. tenuipes and N. serranensis sp. nov are Trans-Andean species, and have even been reported in localities distanced by no more than 60 Km (linear) ( Fig. 2 View FIGURE 2 ), both are clearly differentiable by means of discrete characters, and to a lesser extent by some measurements ( Tables 1, 2, and 3). With individuals of both species in hand, N. serranensis sp. nov. can be easily recognized by its distinctive gray-based ochraceous-buff ventral fur, which is markedly different from the completely white ventral coloration of N. tenuipes ; some specimens of this last species exhibit a pale orange ventral fur, however, basal hairs are not gray as in N. serranensis sp. nov. ( Table 3; Figs. 8A, 8B and 8C View FIGURE 8 ). Also, N. serranensis sp. nov. has broad orange-ochraceous patches on each side of the muzzle (above the mouth) that are absent or slightly pronounced (but narrower) in N. tenuipes ( Figs. 7A and 7B View FIGURE 7 ). Individual hairs covering the dorsal surface of the hindfeet are also useful to differentiate both species; under magnification, most of these hairs appear completely white in N. tenuipes , whereas in N. serranensis sp. nov they are predominately grayish ( Fig. 6 View FIGURE 6 ). In close inspection of the tail, the caudal scales of N. tenuipes appear to be larger than in N. serranensis sp. nov., but this difference is hard to quantify because tails are stretched to varying degrees when skins are stuffed ( Voss et al., 2001). Morphometrically, there is substantial overlapping between the two species in most external measurements ( Tables 1, and 2; Fig. 4 View FIGURE 4 ), however, the holotype of N. serranensis sp. nov. apparently has a longer tail than most of the specimens of N. tenuipes (at least those distributed west of the Magdalena river).
In side-by-side comparisons of the skulls, several differences can be noted between the two species. In dorsal view, the rostrum of N. serranensis sp. nov. is clearly shorter, which morphometrically is reflected by lower values of Rostral Length (RL), Length of Nasals (LN), Length of Incisive Foramen (LIF), and Length of Diastema in the holotype than in most of the specimens of N. tenuipes ( Tables 1, 2; and 3; Figs. 4 View FIGURE 4 , 9A and 9B View FIGURE 9 ). The anterior borders of the nasals are more expanded in N. serranensis sp.nov. than in N. tenuipes . At the level of the interorbital region, the difference is given principally by the supraorbital beads, which are more developed and dorsally projected in N. tenuipes than in N. serranensis sp. nov. Nevertheless, this difference should be interpreted with caution because interorbital beads are less developed in young individuals, as is the case of the holotype of N. serranensis sp. nov. (young adult class II). In lateral view, N. serranensis sp. nov. can be quickly recognized by the convex profile conferred by its globose braincase, while in most specimens of N. tenuipes the profile is flatter ( Figs. 10A and 10B View FIGURE 10 ). From this view is also possible to note a difference in the dorsal surface of the anterior borders of the nasals; in N. tenuipes a subtle hump gives them a hooked appearance, whereas a flatter profile is typical in N. serranensis sp. nov. Posteriorly, at the level of the auditory bullae, there are even more marked differences; the opened ectotympanic ring is distinctive in N. serranensis sp. nov., resulting of the anterodorsal edge of the ectotympanic not being in contact with the mastoid tubercle. Contrary, in N. tenuipes both structures are in contact, which lead to a closed ectotympanic ring ( Fig. 11 View FIGURE 11 ). In N. serranensis sp. nov. the tegmen tympani is exposed above the dorsal surface of the ectotympanic, acting as a membrane that partially covers the ventral portions of the subsquamosal and posglenoid foramina; in N. tenuipes , a non-exposed tegmen tympani leads to a larger posglenoid foramen. In this region, N. serranensis sp. nov. also exhibits a distinctive thicker hamular process of the squamosal than in N. tenuipes , as well as a larger orbicular apophysis of the malleus. With respect to the paraoccipital process, this element is slightly bifurcated in N. serranensis sp. nov, but not in N. tenuipes
In the ventral view of the skull, the first apparent difference between both species involves the incisive foramina, which extend posteriorly almost, but not quite, to the anterior alveolus of M 1 in N. serranensis sp. nov., whereas in N. tenuipes its posterior margins end before ( Figs. 9C and 9D View FIGURE 9 ). Also, on the maxillary of the holotype of N. serranensis sp. nov., adjacent to the posterior margins of the incisive foramina, there are small perforations that are not evident in specimens of N. tenuipes . Dental morphology is another quick way to differentiate between both species; in the maxillary molars of N. serranensis sp. nov, for example, the principal labial cusps (paracone, metacone) are smaller than their lingual counterparts (protocone, hypocone), but in N. tenuipes the labial and lingual cusps are subequal (resulting in a more or less symmetrical occlusal design) ( Fig. 11 View FIGURE 11 ). N. serranensis sp. nov. also has a characteristic M1 with laterally flattened procinculum, contrary to N. tenuipes , in which this tooth is approximately rectangular in outline due to a broad anterocone (not flattened procingulum). The folds of the first upper molars are particularly deep, broad, and bifurcated at their ends in N. serranensis sp. nov, but not in N. tenuipes . In the first species the mesoflexus is divided into a short labial cleft and a large internal fossette that forms a very prominent landmark in the center of the tooth, whereas in the second this element is also divided but the internal fossette is considerably smaller. Finally, in the mandible, there are two useful characters that allow to discriminate between both species: the first is the condylar process, which is notably wide and short in N. serranensis sp. nov, while in N. tenuipes is slightly longer and slender. The second character is the mental foramen, typically larger in N. serranensis sp. nov., and projected laterally, whereas in N. tenuipes is much smaller and is almost dorsally projected.
The enormous gap between the collecting localities of N. serranensis sp. nov. and the known geographical ranges of the other small-bodied species within the genus (implying many physical and environmental barriers), makes impossible to collect them in the same place. Anyway, N. serranensis sp. nov. is easily distinguishable from N. dubosti , N. guianae , N. macedoruizi , N. marajoara , N. minutus , N. musseri , N. paracou , N. vossi , and N. xingu by its unique gray-based ochraceous-buff ventral fur, its broad ochraceous-orange patches in the muzzle, and for being larger in practically all external and craniodental dimensions ( Tables 2, 3, 4, and 5).









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.