
Plicatellopsis bowerbanki ( Vosmaer, 1885 ) comb., 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4755.2.6 |
|
publication LSID |
urn:lsid:zoobank.org:pub:666FEC49-E2D7-4AB1-B89C-3941C0BC1E91 |
|
DOI |
https://doi.org/10.5281/zenodo.3812523 |
|
persistent identifier |
https://treatment.plazi.org/id/2DEB1044-86E4-4281-B597-BAD49B4C6353 |
|
taxon LSID |
lsid:zoobank.org:act:2DEB1044-86E4-4281-B597-BAD49B4C6353 |
|
treatment provided by |
Carolina |
|
scientific name |
Plicatellopsis bowerbanki ( Vosmaer, 1885 ) comb. |
| status |
comb. |
Plicatellopsis bowerbanki ( Vosmaer, 1885) comb. nov.
( Figure 5–6 View FIGURE 5 View FIGURE 6 , Table 4)
urn:lsid:zoobank.org:act:
Original description: Phakellia bowerbanki Vosmaer, 1885: 24 , Pl. I, Fig. 18, Pl. IV, Fig. 7–8, Pl. V, Fig. 45–47.
Synonyms: Cribrochalina ambigua Marenzeller, 1886 , 9, Fig. 1 View FIGURE 1 ., Isodictya dicksonii Fristedt, 1887 , 427–428, Pl. 24, Fig. 14.
Material examined
Norway: BMNH 1910.1.1.1477 “ Phakellia bowerbanki, Willem Barents Expedition, Dr. Vosmaer , portion of Type ” [var. b] ( Portion of lectotype. ZMA. POR.1781) 72.2347N, 22.5E; BMNH 1910.1.1.609.a GoogleMaps “ Phakellia bowerbanki , Type, 12B” [var. b] ( Also portion of lectotype. ZMA. POR.1781); BMNH 10.1.1.610.a “ Phakellia bowerbanki , Type, 12A” [var. α] 72.6013N, 24.95138E. Baffin Bay: CMNI 2018 -0135 (Western Greenland Shelf ) 67.9674N, 59.4847W, 878 m depth; GoogleMaps CMNI 2018 -0136 (Western Greenland Shelf ) 67.9673, 59.4838W, 877 m depth; GoogleMaps CMNI 2018 -0145 (Western Greenland Shelf ) 67.9673N, 59.4840W 877 m depth; GoogleMaps CMNI 2018 -0202 ( Near Pond Inlet, Northern Baffin Island ) 72.8289N, 77.6090W 856 m depth. GoogleMaps Additional collected specimens are noted in Table 4.
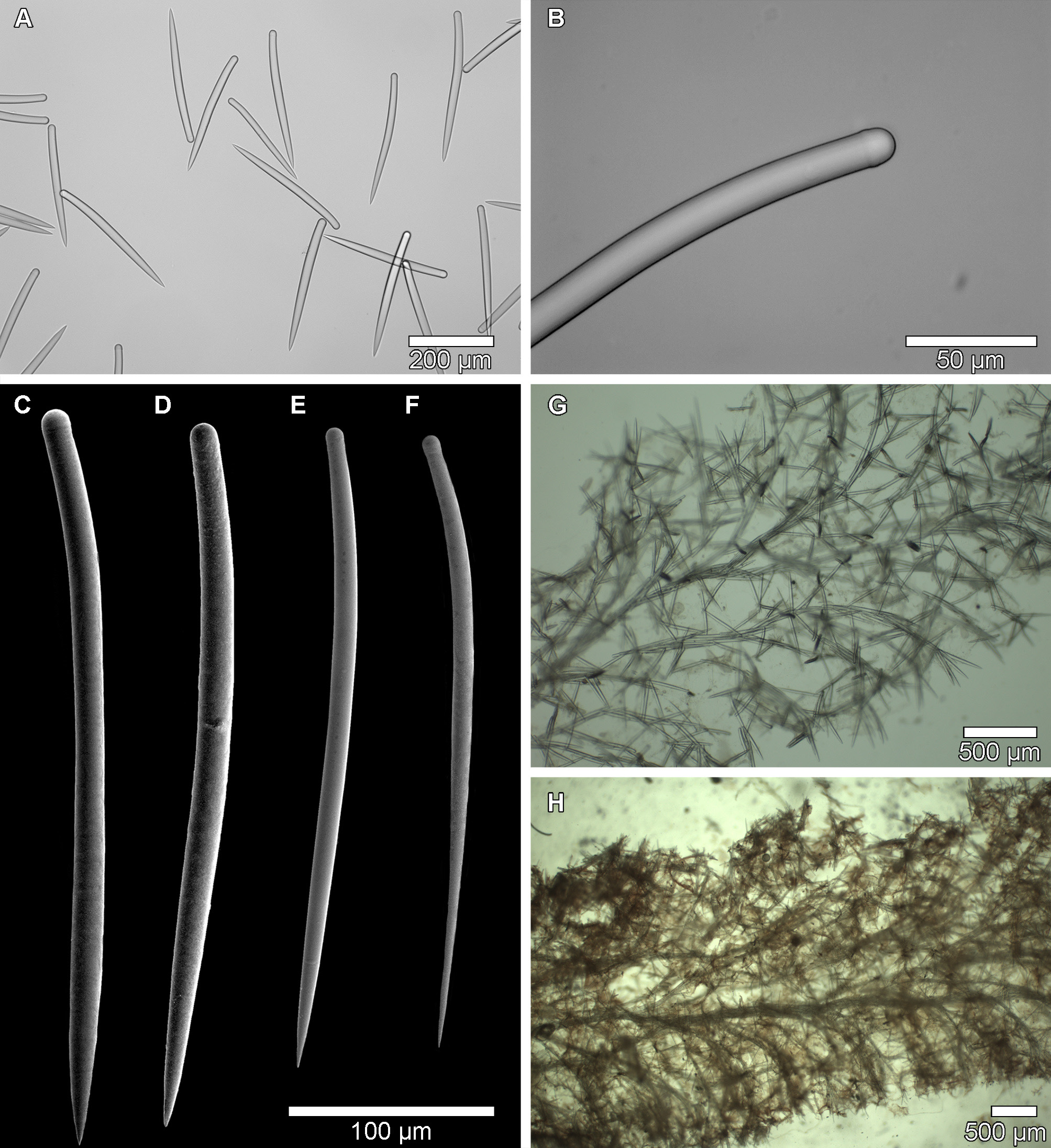
External appearance ( Figure 5 View FIGURE 5 )
Plicatellopsis bowerbanki ( Vosmaer, 1885) comb. nov. is generally a large, vase shaped sponge that forms a funnel at the base ( Fig. 5 View FIGURE 5 A–E). However, the sponge is somewhat polymorphic, and a more fan shape form has been observed that grew to 30 cm wide ( Fig. 5 F View FIGURE 5 ). In some individuals, secondary vases may extend from the distal portions of the primary vase ( Fig. 5 C View FIGURE 5 ). The primary stalk attaches to a hard substrate, usually rock walls, but can grow on dead coral skeletons and pebbles. The inner and outer surfaces have irregularly spaced pores < 1 mm in diameter. In larger specimens, the outer surface is rippled, with ridges and depressions. In larger specimens the raised por- tions appear to form concentric rings or lateral ribs on the underside of the cup ( Fig. 5 C View FIGURE 5 ), but when viewed from above the ridges appear as depressions ( Fig. 5 D, F View FIGURE 5 ). The distal lip of the fan is smoothly curved but in an irregular scalloped manner, and is often frayed both in situ and after collection. Most specimens have large circular holes irregularly placed throughout the sponge body which can include the stem ( Fig. 5 B View FIGURE 5 , D–F, Vosmaer 1885, Pl. 1 Fig. 18). It is unknown if these holes form naturally or are caused by spongivorous predators. However, since no specimens appeared to have predators on or near the sponge body in situ , or after collection, and because the holes are present on specimens throughout the large geographic area in which the sponge was collected, it is suggested that the apertures occur naturally and are thus considered diagnostic of the species.
Spicules ( Figure 6 View FIGURE 6 A–F)
The spicules of Plicatellopsis bowerbanki ( Vosmaer, 1885) comb. nov. are stout, slightly bent tylostyles with very slight tyles. Most spicules look like styles at lower magnifications ( Fig. 6 A View FIGURE 6 ), but the tyles are evident upon closer inspection ( Fig. 6 View FIGURE 6 B–F). The tylostyles can sometimes be separated into two thickness categories, but the thick and thin spicules are consistently similar lengths, and in some specimens the thickness categories are indistinguishable as the widths may overlap. Thinner tylostyles, if present, are not as common and are sometimes difficult to find in spicule preparations due to their inconsistent distribution throughout the sponge tissue. For this reason, thin spicule placement in the skeleton does not appear to be a useful taxonomic character of the species. The skeleton of the lectotype (BMNH 10.1.1.1477 Phakellia bowerbanki Vosmaer, 1885 var. β) was examined here and tylostyles with slight tyles were present, measuring 213– 273 –326 x 8– 13 – 16.5 µm. Only two spicules that were less than 10 µm in thickness were measured from the lectotype. This suggests that the less common thin tylostyles are also present in the specimens from the original description by Vosmaer (1885), but a sufficient number could not be measured as they were not visible in the slide preparation. A specimen from Baffin Bay, CMNI 2018-0202 has tylostyles that are 258– 317 –359 x 14– 18 –21μm, and thinner spicules were not seen. A comparison of spicule sizes for specimens collected is presented in Table 4.
Skeleton ( Figure 6 G, H View FIGURE 6 )
The skeletal architecture of the species consists of dense spicule tracts forming an axial skeleton, which branch out into extra-axial bundles towards the surface, terminating in spicule brushes ( Fig. 6G, H View FIGURE 6 ). The axial and extra-axial spicule tracts consist of between 2–6 spicules. The axial skeleton of tight spicule bundles causes the sponge to have a fibrous texture when cut. When present, the thinner tylostyles are not localized in the skeleton, but rather appear to be loosely placed throughout the sponge tissues ( Fig. 6 G View FIGURE 6 ). The skeleton matches the definition of the genus given by Van Soest (2002) and Burton (1932), except in the surface skeleton where no palisade of smaller spicules is present in the lectotype or in specimens from eastern Canada. In individuals that have a second category of thinner tylostyles, those spicules are not concentrated at the surface like the smaller spicules in other species of the genus, but rather they occur scattered in the choanosomal skeleton without a consistent distribution. The spicules outside of the axial and extra-axial skeletons form a loose, irregularly aligned reticulation of bundles and single spicules.
Genetic data
COI mtDNA sequences were obtained for CMNI 2018-0136 (GenBank MK570857 View Materials ), CMNI 2018-0145 (GenBank- MK570858 View Materials ), CMNI 2018-0202 (Genbank: MK561021 View Materials ), and ARC 81425 (GenBank MK570859 View Materials ). There is no genetic difference in the COI Folmer fragment between specimens from the Gulf of St. Lawrence, Western Greenland Shelf, or Pond Inlet despite differing external morphologies. The 28S gene was sequenced for CMNI 2018-0202 and confirms placement of the species within the Suberitidae , but sequence quality was not sufficiently high to submit to Genbank.
Distribution and ecology
Plicatellopsis bowerbanki ( Vosmaer, 1885) comb. nov. appears to be ubiquitous on the east coast of Canada, extending from the northern tip of Baffin Bay to the northern portions of the Gulf of St. Lawrence. In Canadian waters it is found at depths ranging between 73 and 878 m. Plicatellopsis bowerbanki ( Vosmaer, 1885) comb. nov. is abundant in the Banc-des-Américains Marine Protected Area and the Disko Fan Conservation Area which were recently closed to bottom contact fishing by the Department of Fisheries and Oceans, Canada. Plicatellopsis bowerbanki ( Vosmaer, 1885) comb. nov. has been seen growing on varied substrates, but often attached to harder objects such as dead coral, pebbles and rock walls in otherwise soft sediment environments. The species has also been collected in the waters off Norway, The Faroe Islands, Iceland, Greenland, and in the Barents Sea. ( Burton 1930; Fristedt 1887; Hentschel 1929; Lundbeck 1909; Marenzeller 1886).
Remarks
The original description of Phakellia bowerbanki ( Vosmaer, 1885) stated that placement of the species in the genus was “dubious” as the sponge was lacking “flexible spicules”, likely in reference to vermiform strongyles. Vosmaer argues that Schmidt, Carter, Ridley and others have placed other flabelliform species which lack sinuous spicules in the genus and thus follows this precedent by placing Phakellia bowerbanki var. α and var. β, as well as Phakellia arctica in the genus Phakellia . P. arctica has subsequently been transferred to the genus Axinella (originally Spongia calyciformis ( Lamarck 1814)) , due to the presence of oxeas and styles in the skeleton of that species.
Vosmaer (1885) describes the species as “Great, thin, or funnelshaped [sic]” with strong skeletal fibres. It was also noted that most specimens were torn. Plate I, Fig. 18 ( Vosmaer, 1885) shows a large, red coloured fan-shaped sponge with the characteristic circular aperture, which was also noted in material collected throughout eastern Canada. The deep red or purple colour was not seen in our specimens, although several had a light pink hue after preservation in 95% ethanol. The colour may depend on the method of collection, exposure to air, or the means of preservation, but this remains to be tested.
The original description lists the spicules as acuate ( tr. ac.), and stout. Plate V, Figs. 45–47 ( Vosmaer 1885) clearly show that the spicules for both varieties contain styles, each with a slight bend towards the rounded end. Koltun (1959) describes the spicules as styles, also figured with a slight bend, measuring 218–374 µm. Vosmaer and Koltun both suggest that thinner styles were also present, but were of a length similar to the thicker variety. The original description for the synonymous Cribrochalina ambigua Marenzeller, 1886 shows styles with very slight apical swellings measuring 230–340 µm. Isodictya dicksonii ( Fristedt 1887) from Baffin Bay is also described as a large fan-shaped sponge with smooth styles 300 µm in length. I. dicksonii was listed as a synonym of P. bowerbanki by Hentschel (1929). A sponge listed as “ Phakellia dicksoni (Fristedt) ” by Breitfuss (1912) from the Barents Sea likely represented the same species, although no description was given.
The skeleton of P. bowerbanki ( Vosmaer, 1885) comb. nov. was not described in great detail previously. Vosmaer stated only that the skeletal fibres were strong (var. α) or not much developed (var. β). Koltun described a skeleton consisting of “thick fibres extending along the body and parallel to it”, with “transverse bundles of spicules, as well as individual spicules” (from an English translation of the original Russian manuscript, Koltun 1959). The skeleton of the lectotype ( Fig. 6 H View FIGURE 6 ) shows clearly that the tylostyles form a dense axial skeleton, and the extra-axial skeleton fans outwards towards the surface forming spicule brushes. The section of the lectotype does indeed show a well-developed axial skeleton, contrary to Vosmaer’s original description of var. β. There does not seem to be sufficient difference between the type specimens of varieties α and β for the specimens to be considered separate species.
Phakellia generally has a thick primary skeleton composed of sinuous strongyle or strongyoxea tracts, echinated or connected by styles, strongyles, or strongyloxeas ( Alvarez & Hooper 2002). The genus also has no specialized ectosomal skeleton. The specimens reviewed here do not fit this description as they lack strongyles and strongyoxeas. The skeleton formed of axial and extra-axial tracts, tylostyles and the presence of brushes of spicules in the ectosome fit the description of Plicatellopsis , except that there is no palisade of tylostyles at the surface (Van Soest 2002). This is not the first species identified as Phakellia that has been subsequently transferred to the Suberitidae . Phakellia lobata ( Wilson 1902) , described as having tylostyles has been identified as belonging to the genus Suberites ( Alvarez et al. 1998) .
Phakellia ventilabrum (Linnaeus, 1767) was reportedly collected from the Gulf of St. Lawrence by Whiteaves (1874) and Lambe (1896), but is described by Lambe (1896) as only having sinuous styles (440 x 13 µm and 274 x 3 µm); strongyles were not mentioned. Lambe also described a specimen collected from Davis Strait as P. ventilabrum ( Lambe 1900) but did not give spicule measurements for the specimen. It is likely that, based on the presence of styles only, the specimens identified by Lambe were actually S. cribrosa , but these specimens were not reviewed here.
The World Porifera Database lists six species of Plicatellopsis . Five of the six species occur only in the southern hemisphere. A single species, Plicatellopsis borealis Lehnert & Stone (2017) , was recently described from the Bering Sea. Apart from biogeographic considerations, the North Atlantic species described here is clearly distinct from all others due to the consistently short spicules which are smaller than the tylostyles in other members of the genus ( Table 4). Thinner tylostyles are of a similar length to the thicker tylostyles. Thin tylostyles are therefore not considered to be a distinguishing feature of the species. Thin tylostyles occur in some specimens and are rare in others. As tissue from different body regions was routinely dissolved for spicule preparations, thin tylostyles do not appear to have a taxonomic significance. There is no variation between COI Folmer fragment mtDNA in specimens from Davis Strait and the Gulf of St. Lawrence, though this DNA region is conserved in sponges and may not resolve species-level differences ( Erpenbeck et al. 2006). The presence of stout tylostyles and the skeletal arrangement lacking a dense ectosomal palisade of small spicules are unique to the species. A review of spicule sizes and for other members of the genus were presented by Lehnert & Stone (2017). Plicatellopsis borealis Lehnert & Stone (2017) has large subtylostyles, which exceed the maximum length of spicules found in Plicatellopsis bowerbanki ( Vosmaer, 1885) comb. nov., and the small tylostyles are smaller than the minimum length of spicules in eastern Canadian specimens. The external appearance of P. borealis is similar to Plicatellopsis bowerbanki ( Vosmaer, 1885) comb. nov., where the distal edge of the cup is frayed and irregular, and the pores along the body appear to be distributed similarly in both species. However, Plicatellopsis bowerbanki ( Vosmaer, 1885) comb. nov. often has large, rounded holes along the body of the sponge which are not seen in P. borealis .


| BMNH |
United Kingdom, London, The Natural History Museum [formerly British Museum (Natural History)] |
| ZMA |
ZMA |
| POR |
POR |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Heteroscleromorpha |
|
Order |
|
|
Family |
|
|
Genus |
Plicatellopsis bowerbanki ( Vosmaer, 1885 ) comb.
| Dinn, Curtis, Leys, Sally P., Roussel, Mylène & Méthé, Denise 2020 |
Phakellia bowerbanki Vosmaer, 1885: 24
| Fristedt 1887: 427-428 |
| Marenzeller 1886: 9 |
| Vosmaer 1885: 24 |