
Gracixalus quangi, Rowley & Dau & Nguyen & Cao & Nguyen, 2011
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3098.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03C7132B-FFB4-D95D-BFB1-F99091207251 |
|
treatment provided by |
Felipe |
|
scientific name |
Gracixalus quangi |
| status |
sp. nov. |
Gracixalus quangi View in CoL sp. nov.
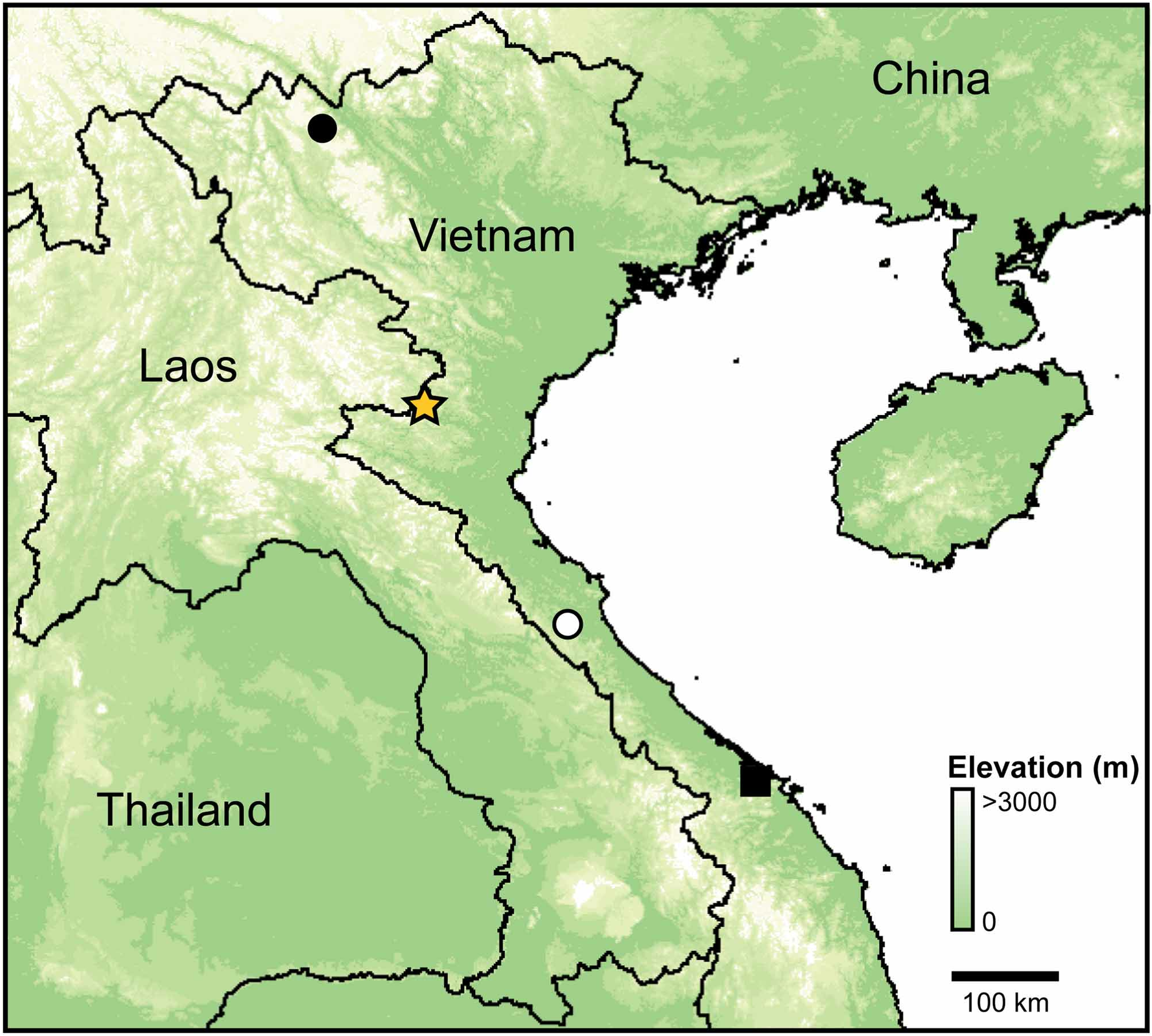
Holotype. AMS R 173410 , adult male, calling on a leaf 1.5 m above rocky seep in a patch of dense, evergreen forest, approximately 10 m from large stream in Pu Hoat Proposed Nature Reserve , Que Phong District, Nghe An Province, Vietnam (19.700 N, 104.738 E, 1,110 m; Figure 1 View FIGURE 1 ). Collected at 19:35 h on 5 June 2010 by Jodi J. L. Rowley, Vinh Quang Dau, Trung Tien Cao, Huong Thi Phung, Tuan Ngoc Le, Luong Thi Nguyen, Trung Danh Hoang, and Thang Thanh Le. GoogleMaps
Paratypes. AMS R 173411, gravid female, collected on leaf adjacent to holotype at the same time as the holotype. AMS R 173412, adult male, calling on mossy twig 1 m above rocky seep, collected at 19:55 h at same locality and date as holotype. AMS R 173413, adult male, calling on leaf 1.5 m above rocky seep, collected at 19:45 h at same locality and date as holotype. AMS R 173414 and NCSM 78276, two adult males collected at 19:40–20:00 h, same date and locality as holotype. AMS R 173415–173416, two adult males, calling, collected at 19:35 and 19:50 h on same date and locality as holotype. AMS R 173417–173418, AMS R 173420, AMS R 173422–173423, NCSM 78277 six adult males, and AMS R 173419, one adult female, collected at 20:00–22:00 h on 6 June 2010 on vegetation above rocky seep approximately 5 m from large, rocky stream (19.700 N, 104.739 E, 1,083 m). VNMN 03000–03010, 11 adult males collected at night on vegetation, adjacent to large rocky stream (19.7613 N, 104.83796 E, 794 m) on 3 July 2010. VNMN 03011, adult female collected at 19:40 h on riparian vegetation above large rocky waterfall (19.769 N, 104.840 E, 1,120 m) on 5 July 2010. All specimens were collected from evergreen forest in Pu Hoat Proposed Nature Reserve, Que Phong District, Nghe An Province, Vietnam. Specimens from June 2010 were collected by Jodi J. L. Rowley, Vinh Quang Dau, Trung Tien Cao, Huong Thi Phung, Tuan Ngoc Le, Luong Thi Nguyen, Trung Danh Hoang, and Thang Thanh Le, and specimens from July 2010 were collected by Vinh Quang Dau, Trung Quang Dau, Hoang Van Bui and Luong Thi Nguyen.
Other material. AMS R 173421, AMS R 173426, two egg clutches collected at ~19:30 h on 5 June 2010 on a leaf 1 m above a rocky seep in a patch of dense, evergreen forest, approximately 10 m from a large stream in Pu Hoat Proposed Nature Reserve, Que Phong District, Nghe An Province, Vietnam (19.700 N, 104.738 E, 1,110 m). AMS R 173424–172425, and AMS R 172427, three egg clutches collected at 20:00–22:00 h on 6 June 2010 on vegetation above a rocky seep approximately 5 m from a large, rocky stream in evergreen forest in Pu Hoat Proposed Nature Reserve, Que Phong District, Nghe An Province, Vietnam (19.700 N, 104.739 E, 1,083 m). Collected by Jodi J. L. Rowley, Vinh Quang Dau, Trung Tien Cao, Huong Thi Phung, Tuan Ngoc Le, Luong Thi Nguyen, Trung Danh Hoang, and Thang Thanh Le.
Etymology. Specific epithet in honour of Vietnamese herpetologist Quang Xuan Hoang, from Vinh University.
Suggested common name. Quang’s Tree Frog (English). Nhái cây Quang (Vietnamese).
Diagnosis. The new species is assigned to the genus Gracixalus by the presence of intercalary cartilage between the terminal and penultimate phalanges of digits, tips of digits expanded into large disks bearing circummarginal grooves, vomerine teeth absent, horizontal pupil, small size (males <25 mm SVL), the presence of large, conical tubercles/spines on the upper eyelid and small white tubercles along the head and cloacal region, triangularly pointed snout, tibia length greater than four times tibia width, translucent skin, and a small number of eggs per clutch laid upon leaves ( Brown & Alcala 1994; Delorme et al. 2005; Duellman & Trueb 1986; Liem 1970). Gracixalus quangi sp. nov. is distinguished from all other rhacophorids in Vietnam and adjacent countries by a combination of (1) small size (males <25 mm SVL), (2) greenish, translucent skin with opaque yellow on anterior surface of thighs and behind insertion of arm, (3) brownish black spots on the flanks and ventral surface of thighs, (4) triangularly pointed snout, and (5) presence of a tibiotarsal projection.
Description of holotype. Body dorsoventrally compressed; head length equal to head width; snout pointed in dorsal view and in profile, projecting beyond margin of the lower jaw; canthus rostralis distinct, sharply angular; loreal region slightly concave; interorbital region convex; nostrils oval, slightly protuberant, closer to tip of snout than eye; pupil horizontal, tympanic rim slightly elevated relative to skin of temporal region, tympanum 38% of eye diameter; pineal ocellus absent; skin not co-ossified to forehead; vomerine teeth absent; choanae small, oval, at margins of roof of mouth; tongue attached anteriorly, very deeply notched posteriorly; pair of distinct, oval vocal sac openings at base of jaw; external subgular vocal sac; slightly raised supratympanic fold extending to just beyond level of axilla. Forelimbs moderately robust, relative length of fingers I <II <IV <III; tips of all fingers with well-developed disks with distinct circummarginal grooves, disks relatively wide compared to finger width (third finger disk 224% third finger width), third finger disk width greater (150%) than tympanum diameter; finger webbing absent; subarticular tubercles prominent, rounded, formula 1, 1, 2, 2; outer palmar tubercle weakly divided into two; accessory palmar tubercles indistinct; nuptial pads present on posteriodorsal surface of Finger I. Relative length of toes I <II <III <V <IV; tips of toes with well-developed disks with distinct circummarginal grooves; disks slightly smaller than those of fingers; webbing formula I 2 + – 2 + II 1 – 3 III 1 - – 3 IV 2 + – 1 - V; subarticular tubercles distinct, rounded, formula 1, 1, 2, 3, 2; inner metatarsal tubercle low, oval, barely distinct, approximately 1 mm long; outer metatarsal tubercle and supernumary tubercles absent. Dorsal surface of head, body, thigh and shank with small, scattered asperities, largest and most concentrated on eyelids. Ventral surface of thighs and belly coarsely granular, chest and throat smooth. Loose skin in gular region. Tarsal fold and dermal ridges absent. Pointed projection at tibiotarsal articulation present, approximately 1 mm long.
Colour of holotype in life. Dorsal surface olive-green, with brighter pale green on the dorsal surface of the upper arms, distal ends of fingers, including toe-disks, thigh and shank ( Figure 2 View FIGURE 2 ). Brown interorbital bar covering eyelids and faint darker “X” across back, joining the interorbital bar anteriorally; faint darker barring on shanks. Bright yellow on the inner surfaces of the thighs, the groin, and in a patch posterior to insertion of arms; yellow spotted with black on the inner thighs. Black supratympanic line underneath supratympannic fold, extending from eye to axilla, covering upper third of tympanum. Opaque, pale turquoise under the supratympanic fold, commencing under the eye and extending posteriorally along the lateral surface of body where it is punctuated by smaller, scattered black spots. A line of large, black spots runs from the supratympanic line to the flanks, separating the turquoise and yellow lower flanks from the olive-green dorsum. The ventral surface of throat, chest and belly are opaque white with translucent pale green margins, the ventral surface of thighs are translucent pale green with small dark brown/black spots, and the ventral surface of the shank are translucent yellowish-green with faint brown barring. The upper arms are ventrally pale, translucent green, with slight pale turquoise speckling at the elbow; the ventral surface of lower arms and hands is pale translucent green with black speckling along the outer margins of lower arm and basal surfaces of Finger III and IV. The pes is ventrally translucent pale green from the tibiotarsal articulation to the inner metatarsal tubercle, including Toe I, and dark blackish green on ventral surfaces of toes II– V. Foot webbing is black in both dorsal and ventral views. Iris bronze with a network of fine black reticulations concentrated around periphery; sclera white. Palpebral membrane with opaque pale turquoise in lower third ( Figure 2C View FIGURE 2 ).
Colour of holotype in preservative. Dorsal surface pale yellowish brown; upper eyelid skin appears black; markings brown ( Figure 3 View FIGURE 3 ). Dorsal surfaces of lower arms, Fingers III and IV, heels and Toes II–V brown. Ventral surfaces pale yellowish cream with dark brown spots on thighs and margins of tibiotarsus. Ventral surfaces of Fingers III and IV, heels, Toes II–V brown. Webbing between toes black. Pale turquoise and yellow on lateral surfaces in life faded to pale cream in preservative.
Measurements. Holotype: SVL 22.8, HDL 8.6, HDW 8.7, SNT 3.6, EYE 3.1, IOD 3.6, TMP 1.2, TEY 0.5, IN 2.6, EN 1.9, TIB 13.3, ML 8.3, PL 10.6, weight in life 0.8 g.
Variation. Measurements of the type series are shown in Table 2. All males have nuptial pads present on Finger I. Dorsal colouration in life varied among individuals and within individuals over time, and may be pale green, dark green, olive-green or pale creamy brown. The number and size of black spots along the lateral surface of the body varied among individuals. The ventral surface of the feet, including webbing, varied temporally from pale greenish or brownish (the same colour as the dorsum) to black. Females are larger and more robust than males.
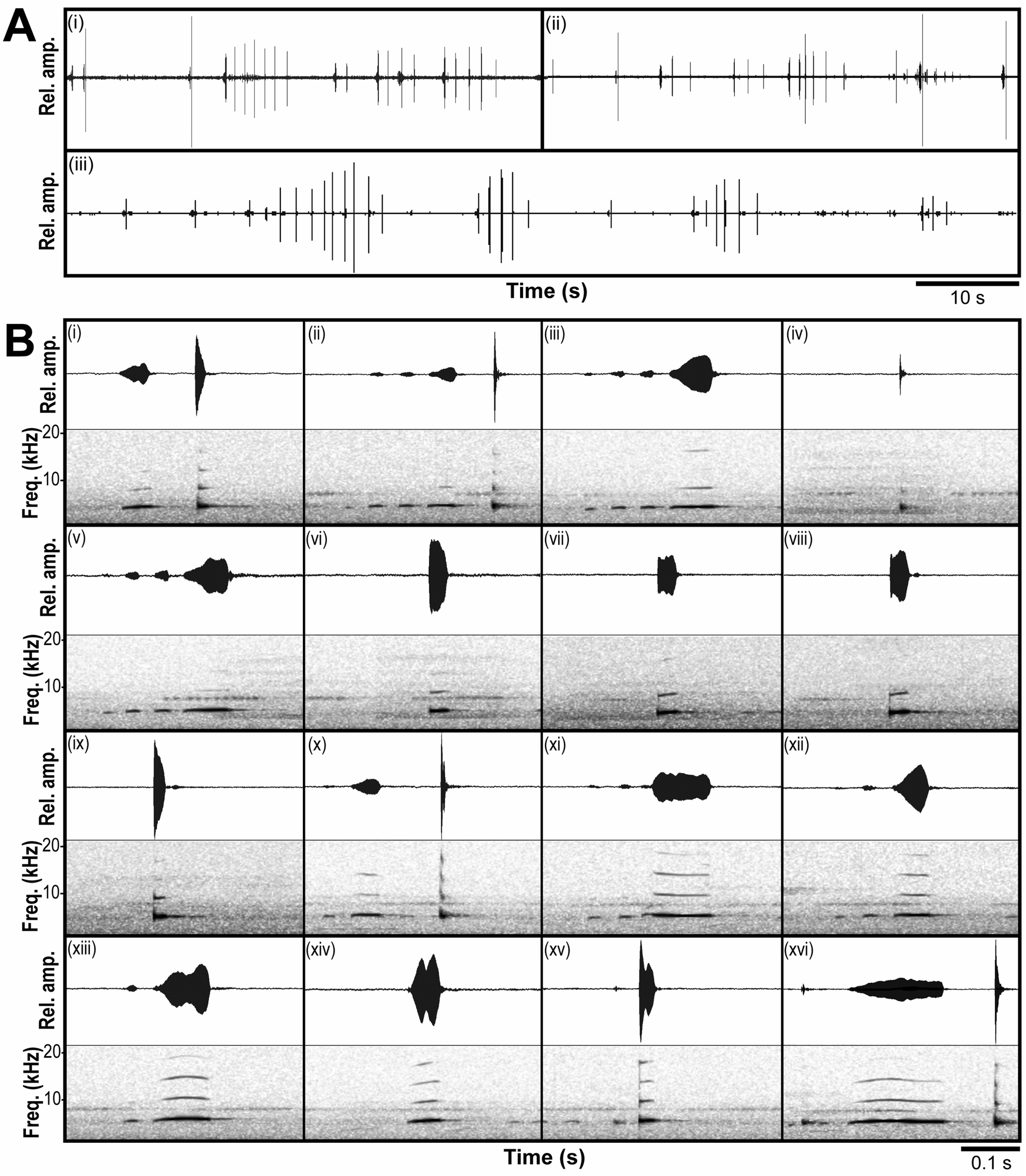
Advertisement call. The male advertisement call of G. quangi sp. nov. consists of a series of highly variable, non-stereotypical call types ( Figure 4 View FIGURE 4 ), ranging from “whistles”, relatively long notes of a relatively narrow frequency band (eg. Figure 4B View FIGURE 4 , call xi) to “clicks”, relatively short notes of a wide frequency band (eg. Figure 4B View FIGURE 4 , call ix). Individual call components were highly variable both within the calling bouts of a single frog ( Figure 4A View FIGURE 4 ) and among individuals. Although some calls were broadly similar in structure (eg. Figure 4B View FIGURE 4 call iii and v, or call vii and iix), no two calls were the same within or among the three individuals recorded. At an ambient temperature of 24.1º C, calls ranged from approximately 10 ms (“clicks”) to 370 ms (calls with “whistle” components). Frequency varied among calls, with calls having a dominant (=fundamental) frequency ranging from 4.1–4.7 kHz, and harmonics present at 8.1–9.6 kHz, 12.0–14.1 kHz, and 16.1–18.8 kHz, depending on the call. Calls were generally frequency modulated and amplitude modulated, but not in a constant or predictable manner ( Figure 4 View FIGURE 4 ). The temporal order of the call types did not appear fixed.
Eggs. As with G. gracilipes and G. supercornutus , G. quangi sp. nov. deposits egg clutches in clumps on leaves overhanging shallow forest pools or puddles ( Figure 5 View FIGURE 5 ). Of six clutches counted, egg number ranged from 7 to 18 (median 11). Eggs are cream coloured and encased in a gelatinous capsule.
Tadpole. Larvae within clutches were observed up to stage 24 of development (Gosner 1960). Recently hatched tadpoles at developmental stage 24 were also observed and collected ( Figures 5–6 View FIGURE 5 View FIGURE 6 ). Measurements of one tadpole at developmental stage 24: total length 10.0 mm, body length 3.3 mm, tail length 6.7 mm, maximum tail height 2.0 mm, tail muscle height 1.1 mm, interorbital distance 1.0 mm. Eyes dorsolateral and oral disk anteroventral. Due to their early developmental stage, tooth rows were not obvious and the oral disk cannot be described adequately. The ‘white spot’ listed as diagnostic for the genus Gracixalus ( Delorme et al. 2005) was not present, which could indicate either that this feature is variable among species within the genus, or that the white spot is present only in more well-developed tadpoles.
Ecology. Gracixalus quangi sp. nov. was observed in small breeding aggregations at several sites throughout Pu Hoat Proposed Nature Reserve in April, June, July and August. Sites where we observed the species ranged from ~ 600–1,300 m elevation, and all sites were in relatively undisturbed, evergreen forest in the Ca and Chu river catchments.
Conservation status. Gracixalus quangi sp. nov. is known only from Pu Hoat Proposed Nature Reserve. The actual distribution of the new species is unknown (currently documented from <15 km 2) but probably extends into adjacent medium to high elevation forested areas in Xuan Lien Nature Reserve in Thanh Hoa Province, in Huaphan Province in Laos, and possibly in nearby Pu Huong Nature Reserve in Nghe An Province. We suggest the species should be considered Data Deficient following IUCN’s Red List categories (IUCN 2001).
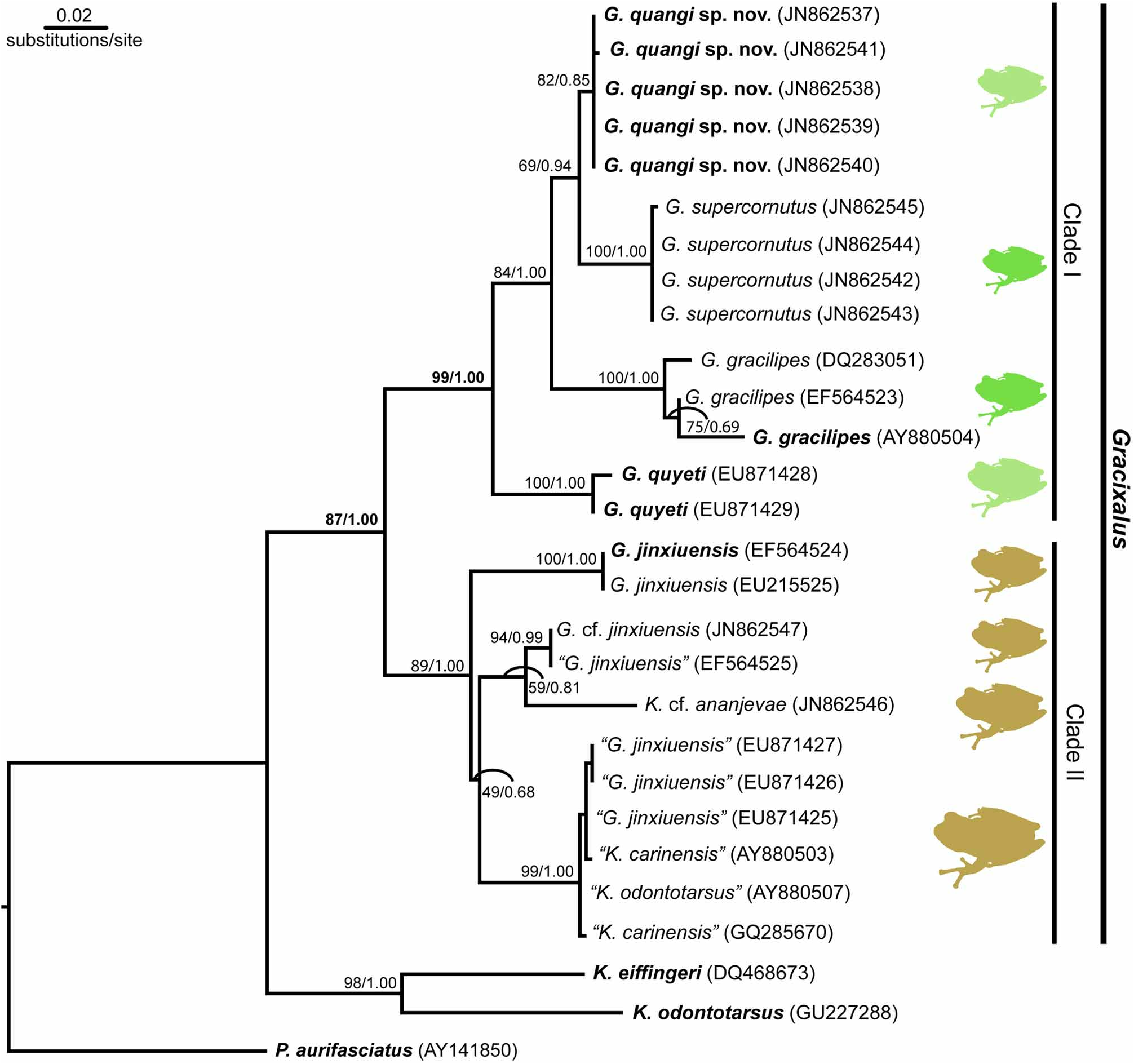
Molecular relationships. Two strongly supported clades were recovered within species currently proposed within the genus Gracixalus ( Figure 7 View FIGURE 7 ). The monophyletic Clade I consists of the type species G. gracilipes and the morphologically similar G. supercornutus , and G. quangi sp. nov., along with G. quyeti (the latter being sister to the previous three species). These four species are relatively small (male SVL <30 mm) with a greenish dorsum ( Figure 8A–E View FIGURE 8 ). Clade II consists of specimens assigned to G. jinxiuensis , G. cf. jinxiuensis , K. cf. ananjevae and K. carinensis (and a sample likely misidentified as K. odontotarsus , as noted previously by Yu et al. 2009). Species in this clade are generally larger in body size with a brownish (never greenish) dorsum and a rounded snout (see Figure 8F–G View FIGURE 8 ).
Further research is required to determine the true identity and phylogenetic relationships of specimens assigned to Gracixalus and related genera. Based on our preliminary phylogenetic analysis, there appears to be multiple lineages of G. jinxiuensis , however some of these lineages may represent misidentified specimens of other species within the group rather than new species. The difficulty in identifying species within this clade is made obvious in the group of samples from close proximity in northern Vietnam (Sa Pa and Fan Si Pan in adjacent Lai Chau and Lao Cai Provinces). Within this group, individuals that are almost certainly the same species (0.2–0.6% sequence divergence at the 16S mtDNA gene fragment) have been identified as three separate species within two genera ( G. jinxiuensis , K. carinensis and K. odontotarsus ). Also noteworthy, the specimen assigned to K. cf. ananjevae from northern central Vietnam fell within Clade II. While K. ananjevae was not compared to any members in this clade in its original description (Matsui & Orlov 2004), the assignment of K. cf. ananjevae to this clade is not surprising given the strong morphological similarities of K. ananjevae to members of this clade, particularly G. jinxiuensis . Known morphological differences between K. ananjevae and G. jinxiuensis are slight (primarily body size), and further detailed morphological and molecular examination of this group is required to assess the validity of known species and identity of new species within clade II. Kurixalus eiffingeri , the type species of the genus Kurixalus , and K. odontotarsus were highly divergent to both clades.
The results of our preliminary analysis provide firm evidence that G. quangi sp. nov. is embedded within the genus Gracixalus (Clade I). This clade receives very high support (99% ML bootstrap support/1.00 Bayesian posterior probability). Molecular differentiation among G. gracilipes , G. supercornutus and G. quangi sp. nov. is relatively low, suggesting recent speciation within the group: uncorrected pairwise distances between the new species and G. gracilipes and G. supercornutus are 4.4–6.2% and 2.2–2.6% respectively. Interspecific variation was low in G. quangi sp. nov. (0.0–0.2%) and G. supercornutus (0.0–0.2%), and higher in G. gracilipes (1.0–3.0; likely a reflection of the c. 75–120 km distance between sampling localities versus <15 km for G. quangi and G. supercornutus sampling localities).
Comparisons. Compared to all other small (adult male SVL <30 mm) species of rhacophorid from Vietnam, Cambodia, Laos and China, the greenish translucent skin, triangularly pointed snout and tibiotarsal projection separates the new species from all but G. gracilipes and G. supercornutus . Buergeria japonica Hallowell 1861 has a brown dorsum, obtusely pointed snout and no tibiotarsal projection; Chiromantis doriae Boulenger 1893 has a pale yellowish brown dorsum, rounded snout and no tibiotarsal projection; Chiromantis laevis Smith 1924 has a rich buff dorsum, obtusely pointed snout and no tibiotarsal projection; Chiromantis nongkhorensis Cochran 1927 has a pale brown dorsum, rounded snout and no tibiotarsal projection; Chiromantis samkosensis Grismer et al. 2007 has a truncate snout and no tibiotarsal projection; Chiromantis vittatus Boulenger 1887 is pale brown, has an obtusely pointed snout and no tibiotarsal projection; Feihyla fuhua Fei et al. 2010b and Feihyla palpebralis Smith 1924 are pale brownish with obtusely pointed snouts and no tibiotarsal projections; Gracixalus jinxiuensis Hu 1978 has a brown dorsum with a rounded snout and no tibiotarsal projection; Gracixalus medogensis Ye and Hu 1984 has a rounded snout and no tibiotarsal projection; Gracixalus quyeti Nguyen et al. 2008 is brownish to moss green with a rounded snout and no tibiotarsal projection; Kurixalus bisacculus Taylor 1962 is dark brown with only a tubercle at the heel; Kurixalus eiffingeri Boettger 1895 has a brown dorsum with a rounded snout and only a tubercle at the heel; Kurixalus idiootocus Kuramoto & Wang 1987 , Kurixalus odontotarsus Ye & Fei 1993 and Kurixalus verrucosus Boulenger 1893 have a brown dorsum with only a tubercle at the heel; Liuixalus hainanus Liu and Wu 2004 , Liuixalus ocellatus Liu & Hu 1973 , and Liuixalus romeri Smith 1953 have a brown dorsum and lack a tibiotarsal projection; Nyctixalus pictus Peters 1871 has a reddish brown dorsum, a truncate snout and no tibiotarsal projection; Philautus abditus Inger et al. 1999 , Philautus cardamonus Ohler et al. 2002 , and Philautus kempii Annandale 1912 have a brown dorsum, rounded snout and no tibiotarsal projection; Philautus maosonensis Bourret 1937 has a brown dorsum, truncated snout and no tibiotarsal projection; Philautus truongsonensis Orlov & Ho 2005 has a brownish dorsum, an obtusely pointed snout, and no tibiotarsal projection; Philautus tytthus Smith 1940 is dark greyish, with a rounded snout and no tibiotarsal projection; Raorchestes gryllus Smith 1924 is variably green, grey or brown in dorsal colour with a tubercle at the heel; Raorchestes longchuanensis Yang & Li 1978 and Raorchestes menglaensis Kou 1990 have a brown dorsum with a rounded snout and no tibiotarsal projection; Raorchestes parvulus Boulenger 1893 has a brownish grey dorsum with a rounded snout and no tibiotarsal projection; Rhacophorus minimus Rao et al. 2006 has a green, non-translucent dorsum, with a bluntly pointed snout and no tibiotarsal projection; Theloderma andersoni Ahl 1927 , Theloderma asperum Boulenger 1886 , Theloderma baibengensis Jiang et al. 2009 , Theloderma lateriticum Bain et al. 2009 , Theloderma rhododiscus Liu & Hu 1962 and Theloderma stellatum Taylor 1962 lack a green dorsum, pointed snout and tibiotarsal projection.
Gracixalus quangi sp. nov. differs from G. gracilipes and G. supercornutus morphologically and molecularly (see below). Compared to G. supercornutus ( Figures 3 View FIGURE 3 , 8C View FIGURE 8 ), G. quangi sp. nov. differs by having a dorsum scattered with small asperities (versus covered in large “spines”), a greenish to brownish green dorsum (versus transparent green with distinct brown markings), brownish black spots on the flanks and ventral surface of thighs present (versus absent), a triangularly pointed snout (versus a more truncate snout forming point only at tip), opaque pale turquoise markings on the lateral surfaces of the head (versus white patch or speckling under the eye to the tympanum), and tibiotarsal projection (versus small tubercle at heel). Compared to G. gracilipes ( Figures 3 View FIGURE 3 , 8D View FIGURE 8 ), G. quangi sp. nov. differs by having a greenish to brownish green dorsum (versus transparent green with brown markings), brownish black spots on flanks and ventral surface of thighs present (versus absent), a triangularly pointed snout (versus a more truncate snout forming point only at tip), opaque pale turquoise markings on the lateral surfaces of the head (versus white patch under the eye to the tympanum), and a tibiotarsal projection present (versus absent or present only as small tubercle). The advertisement calls of G. gracilipes , G. supercornutus , and G. quyeti are unknown.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |