
Pauropsylla jarmilarum, Malenovský & Burckhardt, 2014
|
publication ID |
https://doi.org/ 10.5281/zenodo.5314350 |
|
publication LSID |
lsid:zoobank.org:pub:44343D04-2985-45F4-BA26-4F5C3B481BDAD |
|
DOI |
https://doi.org/10.5281/zenodo.5457251 |
|
persistent identifier |
https://treatment.plazi.org/id/03C787CE-FFA8-8E1F-FE60-CCC7904996A2 |
|
treatment provided by |
Marcus |
|
scientific name |
Pauropsylla jarmilarum |
| status |
sp. nov. |
Pauropsylla jarmilarum View in CoL sp. nov.
( Figs 74–76 View Figs 74–77 , 78–94 View Figs 78–86 View Figs 87–90 View Figs 91–95 )
Type locality. Yemen, central Socotra, Dixam plateau, ca. 1 km NE Shibhon village, limestone cliffs at viewpoint over gorges, 12°29′20″N 53°59′49″E, ca. 680 m a.s.l.
Type material. HOLOTYPE: ♂ ( MMBC, dry-mounted), ‘ YEMEN, SOCOTRA Island / Dixam plateau , view point near / Shibhon village, 15.vi.2012 / 12°29′20″N, 53°59′49″E, 550-680 m / rocks+ Croton socotranus shrubland // inducing pit galls / on Ficus vasta / I. Malenovský leg.’. GoogleMaps PARATYPES: 2 ♂♂ 3 ♀♀, same data as the holotype; GoogleMaps 3 ♂♂, 7 fifth instar and 6 fourth instar immatures, north-eastern Socotra , Homhil protected area , surroundings of Ain Tsahrin spring , 12°34′12″N 54°18′30″E, 435 m, 11.vi.2012, on Ficus vasta , I. Malenovský leg.; GoogleMaps 2 ♂♂ 3 ♀♀, north-eastern Socotra , Kazazhan area , 12°33′48″N 54°19′48″E, 540 m, 10.vi.2012, limestone cliffs, in sink holes , on Ficus vasta , I. Malenovský leg. ( MMBC, NHMB, NMPC; dry- and slide-mounted). GoogleMaps
Description. Adult. Coloration ( Figs 74 View Figs 74–77 , 91 View Figs 91–95 ). Head and thorax in males almost uniformly dark brown to black, in females vertex laterally, mesopraescutum in posterior two thirds and mesoscutum (except for four dorsal dark brown stripes in some specimens) lighter ochreous brown. Antenna basally (segments 1–3 except for apex of segment 3) pale yellow, gradually becoming darker towards apex (apices of segments usually slightly darker): central portion (apex of segment 3 to segment 6) infuscate light brown, segment 7 and base of segment 8 dark brown, apical two thirds of segment 8 and entire segments 9 and 10 black. Rostrum basally pale yellow, apically dark brown to black. Legs uniformly pale yellow except for second tarsal segments of all legs (in both sexes) and metafemur (particularly in males) which are brownish. Forewing membrane hyaline, veins uniformly pale yellow, costal and anal veins basally brownish ( Fig. 76 View Figs 74–77 ). Hind wing hyaline. Abdomen in males with tergites 1–2 light green-yellow, tergites 3–8 dark brown to black, sternites and terminalia green-yellow, first visible sternite basally and male subgenital plate brownish; in females almost uniformly light green-yellow including terminalia.
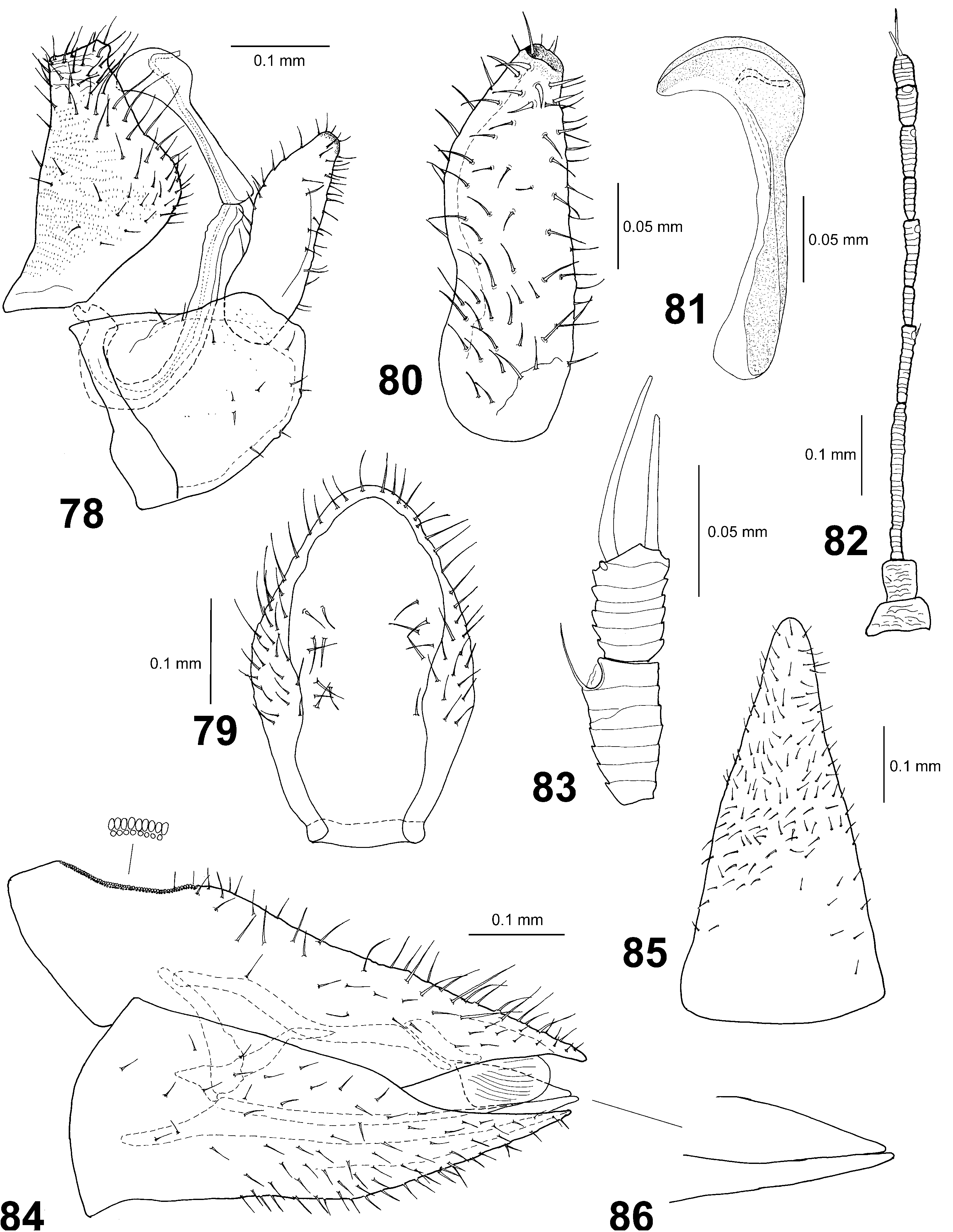
Structure. Integument with fine microsculpture, shiny, covered with very short setae. Head ( Fig. 75 View Figs 74–77 ) with vertex rounded down in front, dorsally elevated on sides, lateral ocelli lying on tubercles above the plane of vertex; median suture of vertex completely developed. Frons narrowly elliptic. Genae rounded, only very weakly bulging laterally below eyes. Eyes prominent, hemispherical. Antenna slender, with 10 segments, segments hardly narrowing to apex, segment 3 longest ( Fig. 82 View Figs 78–86 ); single oval rhinarium subapically on segments 4, 6, 8 and 9; rhinaria not associated with any other conspicuous sensilla; segment 10 with terminal setae subequal, the longer seta ca. 1.3 times longer than segment 10, shorter seta about as long as segment 10 ( Fig. 83 View Figs 78–86 ). Clypeus irregularly hemispherical, with few short setae at apex; rostrum lacking conspicuous setae. Fore trochanter with 3–4 long fine setae, lacking strong ventroapical spur. Metacoxa with well-developed meracanthus. Metatibia elongate, with three (one outer, two inner) dark sclerotised spurs and 10 tightly packed smaller and finer but still stout, unsclerotised setae apically. Metabasitarsus lacking apical spurs, about as long as or slightly shorter than apical tarsal segment. Forewing ( Fig. 76 View Figs 74–77 ) broadest in apical third, with narrowly rounded apex in cell m 1; R+M+Cu stem trifurcating, common M+Cu stem absent; R S vein convex; m 1 cell relatively small, distinctly smaller than cu 1 cell; forewing membrane lacking surface spinules, with radular spinules arranged in narrow triangular fields in cells m 1, m 2 and cu 1; fore wing veins only with minute, inconspicuous setae. Male subgenital plate subglobular, with slightly sinuate dorsal margin and few sparse short setae postero-ventrally ( Fig. 78 View Figs 78–86 ). Male proctiger flask-shaped, medially with well-developed, broadly rounded posterior expansions ( Fig. 78 View Figs 78–86 ); inner surface of each expansion bearing 6–9 fine setae arranged in irregular dorso-ventral line ( Fig. 79 View Figs 78–86 ). Paramere, in lateral view, with nearly straight posterior margin, anterior margin weakly S-shaped: constricted in basal third, convex medially and gradually narrowing in apical third; apex narrowly truncate, well-sclerotised, bearing small tooth oriented towards front; inner face covered with fine setae, subapically with five stouter setae; another row of six stouter setae situated along anterior margin medially ( Fig. 80 View Figs 78–86 ). Distal segment of aedeagus with simple, broadly sickle-shaped apical dilation, ductus ejaculatorius relatively long and sinuate ( Fig. 81 View Figs 78–86 ). Female terminalia very long, covered with numerous short to moderately long setae; proctiger, in lateral view, with dorsal margin straight, apex acutely pointed; circumanal pore ring elliptic with two contiguous rows of pores; subgenital plate, in lateral view, regularly slightly convex ventrally, with long, narrow apical extension, pointed at apex ( Fig. 84 View Figs 78–86 ), in ventral view, narrowly triangular, narrowly rounded at apex ( Fig. 85 View Figs 78–86 ); dorsal and ventral valvulae smooth, lacking any teeth ( Fig. 86 View Figs 78–86 ).
Measurements (in mm). Males (n = 2): HW 0.58, AL 0.71–0.76, WL 1.92–1.94, WW 0.91–0.92, TL 0.71–0.72, MPL 0.26–0.27, PL 0.21, AEL 0.18. Females (n = 2): HW 0.59–0.60, AL 0.73, WL 1.98–2.15, WW 0.95–1.00, TL 0.71–0.72, FPL 0.51–0.53, SL 0.42–0.50. Ratios: AL/HW 1.22–1.31, WL/HW 3.31–3.58, WL/WW 2.08–2.15, TL/HW 1.18–1.24, MPL/HW 0.45–0.47, FPL/HW 0.86–0.88, FPL/SL 1.06–1.21.
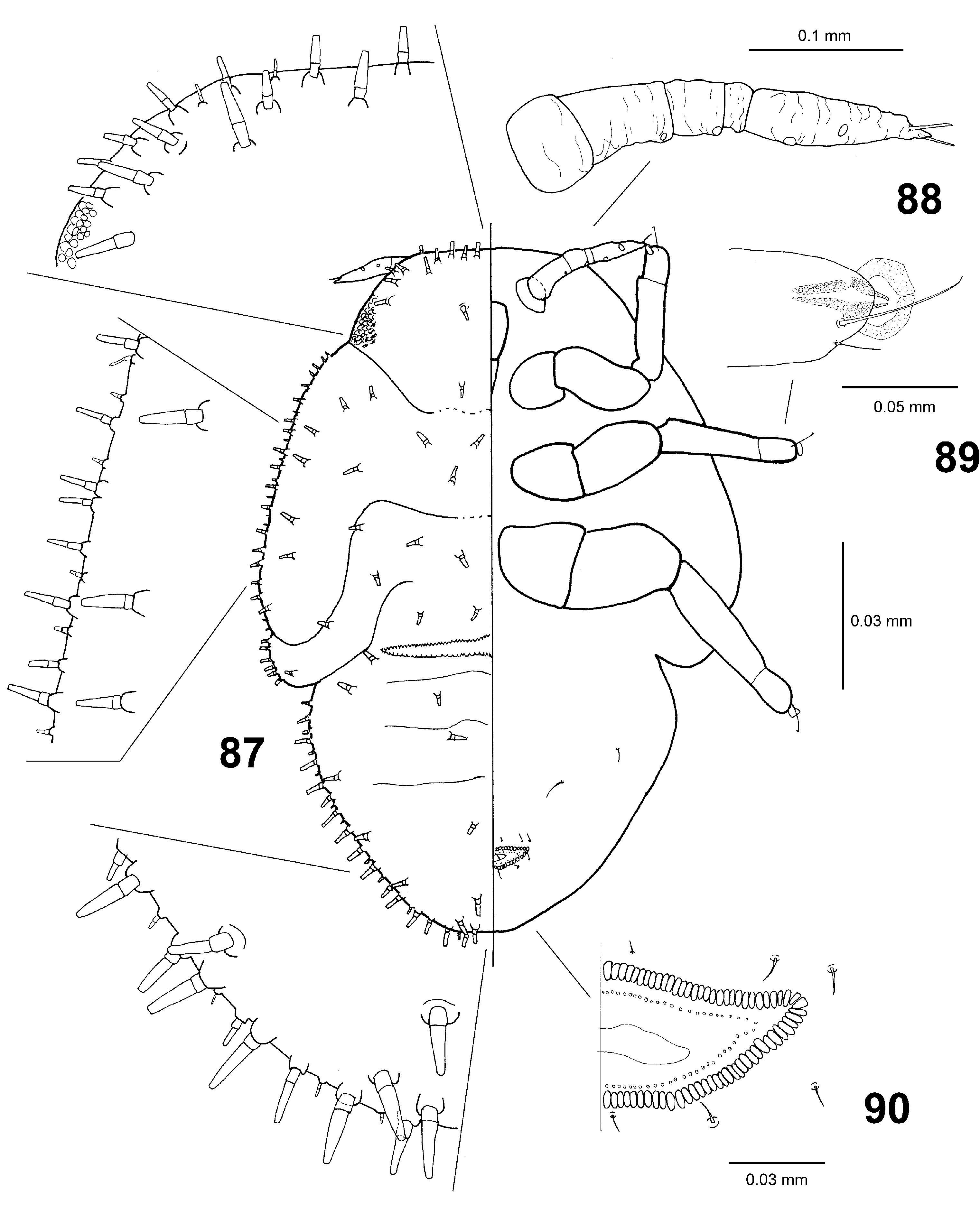
Fifth instar immature ( Fig. 87 View Figs 87–90 ). Uniformly pale yellow-white. Body oval, elongate. Body margin bearing many slender trucate sectasetae arranged in three rows; outer row with alternating large and small sectasetae; dorsum of cephalothorax and caudal plate with sparse similar, large, truncate sectasetae. Antenna divided into five or six segments, bearing four rhinaria ( Fig. 88 View Figs 87–90 ). Cephalothorax partly indistinctly subdivided; posterior margin of cephalothorax and anterior margin of caudal plate finely serrate. Forewing pad with humeral lobe slightly surpassing posterior eye margin. Anus ventral; outer circumanal pore ring consisting of one row of pores ( Fig. 90 View Figs 87–90 ).
Measurements (n = 6; in mm). BL 1.30–1.42, BW 0.90–1.02, AL 0.29–0.33, FL 0.58–0.65, TL 0.34–0.38, CPL 0.50–0.62, CPW 0.69–0.74, CRW 0.12–0.13. Ratios: BL/BW 1.35–1.48, AL/FL 0.49–0.55, CL/CW 0.68–0.83, CRW/CPW 0.16–0.18.
Differential diagnosis. Pauropsylla Rübsaamen, 1899 currently includes 23 valid species from the Afrotropical, Oriental and Palaearctic Regions ( MATHUR 1975, HOLLIS 1984, LI 2011, OUVRARD 2014). The adults of P. jarmilarum sp. nov. can be differentiated from all previously described species by the following combination of characters: median suture of vertex completely developed; antenna 10-segmented, with a single rhinarium each on segments 4, 6, 8 and 9, lacking bifid sensilla; fore trochanter lacking a strong ventroapical spur; metatibia with 2+1 sclerotised apical spurs; forewing relatively narrow with apex narrowly rounded; vein R+M+Cu of forewing trifurcating, common stem M+Cu absent; male proctiger flask-shaped, with moderately developed posterior lobes bearing relatively few fine setae on their inner side; paramere shape; apical segment of aedeagus simply sickle-shaped; female terminalia very long, with subgenital plate narrowly rounded at apex.
The fifth instar immature of P. jarmilarum sp. nov. resembles P. willcocksi Dçbski, 1918 , widely distributed in northern Africa and Arabia, from which it differs in slightly narrower body shape (BL/BW = 1.3 in P. willcocksi ; HOLLIS 1984) and smaller number of marginal sectasetae.Adult P. willcocksi have a strong ventroapical spur on the fore trochanter, numerous peg-like setae on inner side of the lobes of the male proctiger, a strongly curved, sickle-shaped paramere, and female subgenital plate with a ventral transverse groove in apical third ( HOLLIS 1984).
The shape of the male and female terminalia is reminiscent of four species from India, namely P. depressa Crawford, 1912 , P.ficicola Kieffer, 1905 , P. purpurescens Mathur, 1975 , and P. reticulata Mathur, 1975 which are generally larger than P. jarmilarum sp. nov., have forewings with a distinct vein M+Cu, and shorter female terminalia; P. depressa and P. purpurescens also differ in the absence of the median suture of vertex and a more truncate forewing apex and the lack of sectasetae on dorsum of the fifth instar immatures (sectasetae are missing completely in P. depressa ); P. ficicola differs in the concave vein R S and a large m 1 cell of forewing and different number of sclerotised apical spurs on metatibia. Finally, P. reticulata differs in broader and apically truncate forewing and 2+2 apical spurs on the metatibia ( MATHUR 1975).
The Oriental Pauropsylla triozoptera Crawford, 1913 resembles P. jarmilarum sp. nov. in the absence of M+Cu and the long female terminalia but differs in broader forewing and the shape of the paramere which bears a basal anterior lobe, as well as in a number of immature characters ( YANG 1984, as Sympauropsylla triozoptera ; LI 2011, as Neotrioza triozoptera ; YANG et al. 2013).
Etymology. Derived from the Czech feminine personal name ‘Jarmila’. Named after Jarmila Malenovská and Jarmila Jurová, mother and grandmother of the first author.
Host plant. Ficus vasta Forssk. (Moraceae) .
Biology. Immatures induce small pit galls on lower surface of the leaves of the host plant. The galls are circular depressions, ca. 2 mm large in diameter, appearing as light green nipples on the above surface of the leaves. There can be up to ca. 60 pit galls per leaf scattered over the leaf blade; individual galls are situated more or less close to larger veins ( Figs 92–94 View Figs 91–95 ).
Occurrence in Socotra. Found at middle elevations on limestone plateaus in central and north-eastern Socotra, together with the host plant in moderately humid microhabitats on limestone cliffs and boulders, associated often with sink holes and terraces ( Figs 77 View Figs 74–77 , 95 View Figs 91–95 ).
Distribution. So far only known from the Socotra Island. The host plant is widely distributed in southern parts of Arabian Peninsula and mainland eastern Africa ( HASSLER 2014).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |