
Echinoderes sensibilis Adrianov, Murakami & Shirayama, 2002
|
publication ID |
https://doi.org/ 10.5852/ejt.2020.730.1197 |
|
publication LSID |
lsid:zoobank.org:pub:857A9432-9083-46B3-B0BF-B34D619EB350 |
|
DOI |
https://doi.org/10.5281/zenodo.4420029 |
|
persistent identifier |
https://treatment.plazi.org/id/03C79270-FF89-572C-B2D6-FC50190BF998 |
|
treatment provided by |
Plazi |
|
scientific name |
Echinoderes sensibilis Adrianov, Murakami & Shirayama, 2002 |
| status |
|
Echinoderes sensibilis Adrianov, Murakami & Shirayama, 2002
Figs 11–15 View Fig View Fig View Fig View Fig View Fig ; Tables 11–12 View Table 11
Emended diagnosis
Echinoderes with short middorsal spines on segments 4 to 8, and lateroventral spines on segments 6 to 9; middorsal spines on posterior segments barely or just reaching the posterior margin of their respective segments. Tubes present in lateroventral positions on segments 2 and 5, and in laterodorsal positions on 10. Minute glandular cell outlets type 2 in midlateral positions on segments 8 and 9. Tergal extensions of segment 11 short, pointed and well-spaced; sternal extensions short, with ventrolateral seta-like tuft of extended fringe tips. Females with ventromedial female papillae resembling glandular cell outlets type 2 on segments 6 to 8; intracuticular substructure of female papillae crescentic on segment 6, and tubular on segments 7 and 8.
Material examined
Holotype
JAPAN • ♂; Honshu Island , Tanabe Bay ; 33°42′12″ N, 135°22′54″ E; 0 m b.s.l.; Mar. 2001; A. Adrianov, C. Murakami and Y. Shirayama leg.; calcareous red algae ( Corallina pilulifera ) in tidal pool; SMBL CM-A-JAP-E04 . Specimen mounted for LM. GoogleMaps
Paratypes
JAPAN • 3 ♂♂; same collection data as for holotype; SMBL CM-A-JAP-E01 , CM-A-JAP-E05 and CM-A-JAP-E08 GoogleMaps .
Specimens mounted for LM. One specimen was incorrectly identified as female in the original description.
Additional material
JAPAN • 1 ♀; Honshu Island , Tanabe Bay ; 33°41′31″ N, 135°20′10″ E; 0 m b.s.l.; 12 Sep. 2012; H. Yamasaki leg.; detritus and mixed sediment in tidal pool; GenBank: LC557098 View Materials ; ICHUM-6127 GoogleMaps .
DNA was extracted from this specimen, and cytochrome oxidase subunit 1 was sequenced. The cuticle of the specimen was subsequently recovered and mounted for LM ( ICHUM- 6127).
JAPAN • 3 ♂♂; same collection data as specimen above; personal reference collection of the last author. Specimens mounted for SEM.
Additional specimens from the same locality were collected by C.Y. Chang, mounted for SEM, and stored in the personal reference collection of the first author.
JAPAN • 1 ♀; Okinawa, Sesoko Station ; 26°39′00″ N, 127°51′22″ E; 0 m b.s.l.; 10 Nov. 2016; S. Fujimoto leg.; coarse sand in tidal pool; NHMD-115257 . Specimen mounted for LM GoogleMaps .
REPUBLIC OF KOREA • 2 ♂♂; Korean west coast, Wido Island ; 35°36′52″ N, 126°17′01″ E; 0 m b.s.l.; 31 Feb. 2000; J. Lee and Y.H. Song leg.; intertidal macroalgae; NHMD-664200 , NHMD-664201 GoogleMaps • 1 ♀, 1 ♂; Jeju Island , Gimnyeong ; 33°33′33″ N, 126°45′16″ E; 0 m b.s.l.; 1 Mar. 2000; C.Y. Chang, J. Lee and Y.H. Song leg.; intertidal macroalgae; NHMD-664202 , NHMD-664203 GoogleMaps • 1 ♂; Jeju Island , Udo Islet ; 33°31′03″ N, 126°57′40″ E; 0 m b.s.l.; 3 May 2000; C.Y. Chang and J. Lee leg.; intertidal macroalgae; NHMD-664204 GoogleMaps .
All Korean specimens mounted for LM.
HAWAII • 4 ♀♀, 6♂♂; Oahu, Kaneohe Bay, lagoon at Coconut Island , St. MVS-0224-HI-01 (= workshop St. KANM005); 21°26′09″ N, 157°47′18″ W; 0,5 m b.s.l.; 22 May 2017; M. V. Sørensen leg.; subtidal black mud, rich in organic matter; GenBank: MT999943 View Materials ; NHMD-662049 to 662058 GoogleMaps .
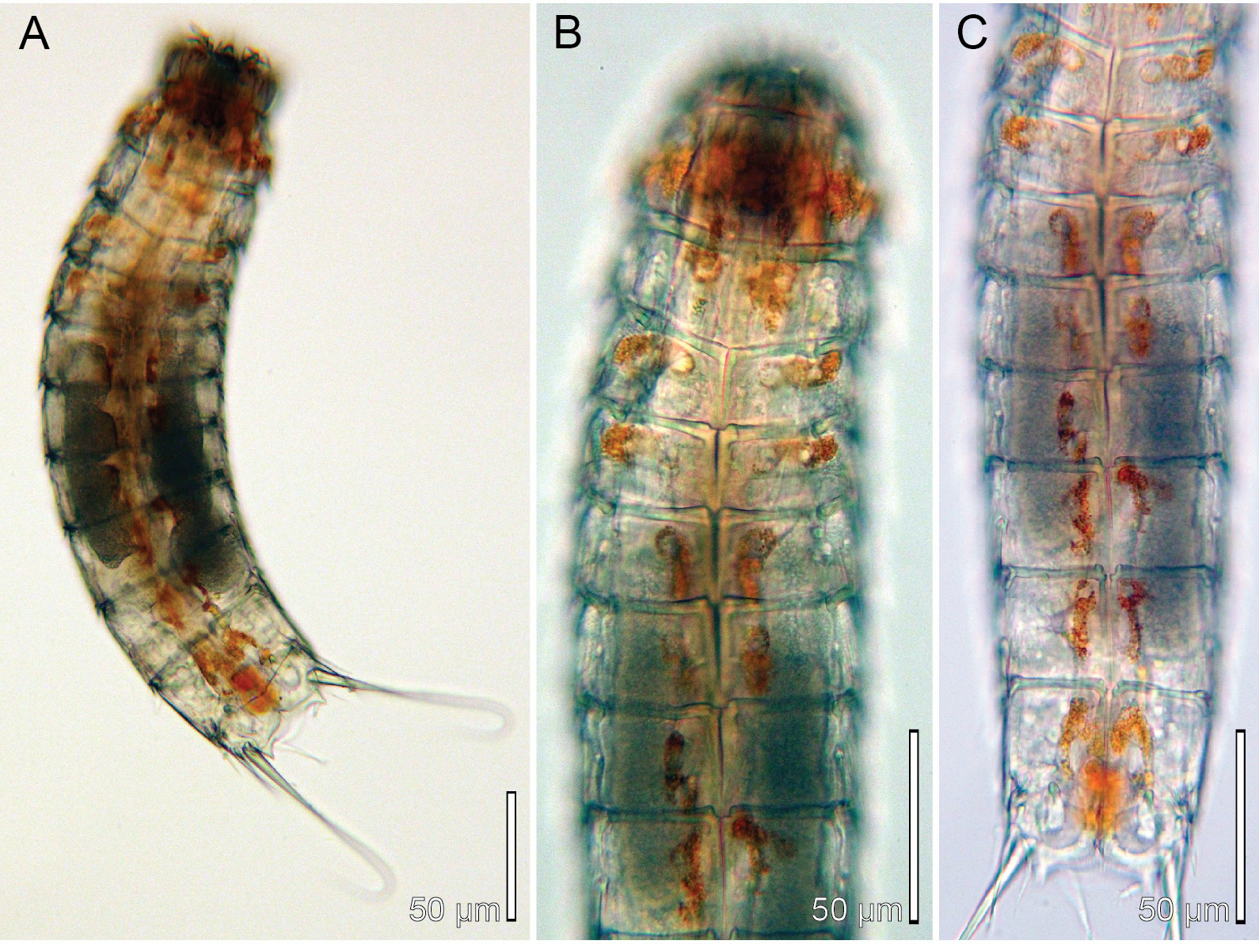
Specimens were mounted for LM. Additional 5 ♀♀, 5 ♂♂ collected at the same station were mounted for SEM and stored in the first author’s personal reference collection. One specimen was picked up for molecular barcoding and a 625 bp long cytochrome oxidase subunit 1 sequence was deposited in GenBank under accession number MT999943 View Materials ; the cuticle could not be recovered afterwards, but the specimen used for DNA extraction is shown on Fig. 11 View Fig .
HAWAII • 6 ♀♀, 4 ♂♂; Oahu, Kaneohe Bay, He’ia Fish Pond , St. MVS-0229-HI-06 (= St. KANM034); 21°26′05″ N, 157°48′19″ W; 3 m b.s.l.; 24 May 2017; workshop participants leg.; subtidal black mud, very rich in organic matter; NHMD-662059 to 662068 GoogleMaps .
Specimens were mounted for LM. Additional 7 ♀♀, 3 ♂♂ collected at the same station were mounted for SEM and stored in the first author’s personal reference collection.
See Table 1 View Table 1 for an overview.
Redescription
Since the present redescription adds several new features to the species, including information on head structures, observations of live material, and significant diagnostic traits, and since the distributional range of the species is extended considerably, we are providing a full description for the species. As much information as possible is based on observations of the type material, but the specimens are partly deteriorated, hence structures such as sensory spots could no longer be observed. Additional information is primary based on examination of topotypes, supplemented with information from Okinawa and Hawaii populations that morphologically agreed completely with the topotypes.
Adults with head, neck and eleven trunk segments ( Figs 11–13 View Fig View Fig View Fig , 14A View Fig , 15A View Fig ). Glandular cells appear brown-red in live specimens, giving the species a conspicuous longitudinal colour pattern along its trunk ( Fig. 11 View Fig ). The colouring fades, and eventually disappears, when the animal is fixed. The trunk appears stout, with nearly equally broad sternal plates on segments 6 to 10 ( Figs 11A, C View Fig , 14A View Fig ). Segments 1 to 4 were often strongly contracted in fixed specimens. Lateral terminal spines slender, from 40% to 55% of trunk length. For complete overview of measurements and dimensions, see Table 11 View Table 11 . Secondary pectinate fringe present near anterior segment margin on segments 2 to 10, but it is usually covered by the preceding segment. Distribution of cuticular structures, i.e., sensory spots, glandular cell outlets, spines and tubes, is summarized in Table 12.
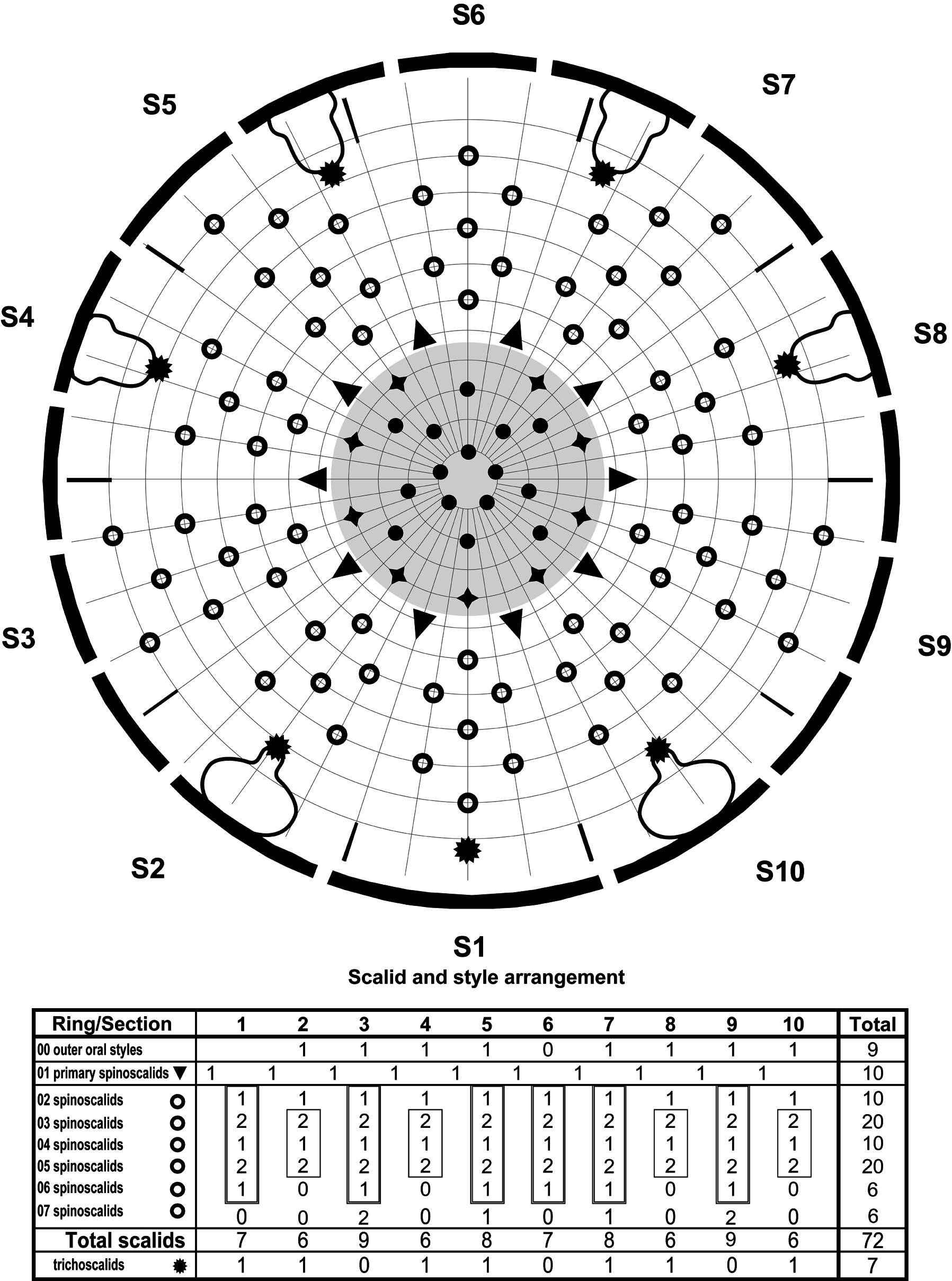
The head consists of a retractable mouth cone and an introvert ( Figs 13 View Fig , 15 View Fig B–C). Three rings of inner oral styles present, with five styles in each ring ( Fig. 13 View Fig ). The external mouth cone armature consists of nine outer oral styles; bases of outer oral styles each with a V-shaped row of short fringe tips and a central bristle of much longer fringe tips ( Fig. 15B View Fig ). The introvert sectors are defined by the ten primary spinoscalids in Ring 01 ( Figs 13 View Fig , 15C View Fig ). Each primary spinoscalid consists of a basal sheath and a distal end piece with a blunt tip. The sheaths have a well-defined basal, transverse fringe, and a less distinct fringe more distally on their exterior surfaces; fringe tips of the distal fringe attach along a longitudinal line and alternatingly projects to the left and to the right (see inset Fig. 4B View Fig for a similar morphology in a different species). End pieces are smooth and flexible. Rings 02 and 04 have 10 spinoscalids and Rings 03 and 05 have 20. All spinoscalids in these rings are well-developed, and consist of a basal sheath and a pointed end piece ( Fig. 15C View Fig ). The basal sheaths terminate into fine, fringed margins in spinoscalids of Rings 02 to 05, and those of Rings 03 to 05 have in addition a basal median spike. A ring of short fringes extend around the introvert in between spinoscalid Rings 04 and 05. Ring 06 has only six spinoscalids, located in sectors 1, 3, 5, 6, 7, and 9 ( Fig. 13 View Fig ); Ring 06 spinoscalids resemble those in preceding sectors, but without a distinct differentiation into sheath and end piece. Ring 07 also has 6 spinoscalids, located as pairs in sectors 3 and 9, and unpaired but laterally displaced in sectors 5 and 7 (trichoscalids are taking up the space in the opposite side of each sector); Ring 07 spinoscalids resemble those in preceding sector.
Described sector-wise ( Fig. 13 View Fig ), sectors 1 and 6 are similar, having spinoscalids arranged as two double diamonds. Sectors 2, 4, 8 and 10 all have spinoscalids arranged as a quincunx, located in between a medial anterior spinoscalid (Ring 02) and a trichoscalid plate. Sectors 3 and 9 have spinoscalids forming double diamonds anterior to a pair of spinoscalids. Sectors 5 and 7 also have spinoscalids forming double diamonds, but anterior to an unpaired, lateral spinoscalid ( Fig. 13 View Fig ).
Regular trichoscalids with trichoscalid plates are present in sectors 2, 4, 5, 7, 8, and 10. In addition, a single trichoscalid without trichoscalid plate is present in sector 1 ( Figs 13 View Fig , 14H View Fig , 15C View Fig ). The trichoscalid has the typical furry appearance ( Figs 14H View Fig , 15C View Fig ), but basally it also has a series of more well-organized pointed denticles.
The neck has 16 placids, measuring 18 µm in length. The midventral placid is broadest, measuring 15 µm in width at its base, whereas all other are narrower, measuring 11 µm in width at their bases. The trichoscalid plates are well-developed; subdorsal and laterodorsal ones are narrow and elongated, and ventromedial ones broadly oval.
Segment 1 consists of a complete cuticular ring. Sensory spots are located near the anterior margin in subdorsal and laterodorsal positions, and slightly more posterior in sublateral and ventromedial positions ( Figs 12 View Fig A–B, 14B, 15D–E); sensory spots are large, and rounded to oval, with numerous micropapillae, two pores, and often a cilium emerging from one of the pores. Glandular cell outlets type 1 present in middorsal and lateroventral positions ( Figs 12 View Fig A–B, 14B–C, 15D–E); all glandular cells with type 1 outlets on this and following segments with conspicuous brown-red colouring that gets lost during fixation ( Fig. 11 View Fig A–B). Dorsal and lateral sides, and posterior half of ventral side, with scattered cuticular hairs emerging through rounded perforation sites. The posterior segment margin is straight around the segment, terminating into a pectinate fringe with short, uniform fringe tips.
Segment 2 consists of a complete cuticular ring. Pachycyclus of the anterior segment margin is of medium thickness and not interrupted ( Figs 11B View Fig , 14 View Fig A–C). Sensory spots are located in middorsal (but slightly laterally displaced), laterodorsal (twin pair) and ventromedial positions ( Figs 12 View Fig A–B, 15D–E); sensory spots on this and all following segments as on segment 1, but slightly smaller. Glandular cell outlets type 1 present in middorsal and ventromedial positions, and quite well-developed tubes present in lateroventral positions ( Figs 12 View Fig A–B, 14B–C, 15D–E). The segment is densely covered with bracteate hairs; hair cover slightly thinner between ventromedial sensory spots. The posterior segment margin is nearly straight; pectinate fringe from middorsal to midlateral positions with short fringe tips, as on segment 1; fringe tips from midlateral to ventromedial positions conspicuously longer, and then again very short between ventromedial positions.
Segment 3, and remaining segments, consisting of one tergal and two sternal plates. Pachycyclus of the anterior segment margin of medium thickness, and interrupted only at tergosternal junctions ( Figs 11 View Fig B–C, 14C). Sensory spots present in subdorsal, laterodorsal and sublateral positions ( Fig. 12 View Fig A– B). Glandular cell outlets type 1 present in middorsal and ventromedial positions ( Figs 12 View Fig A–B, 14B–C). Bracteate cuticular hairs are densely covering the segment from middorsal to ventromedial positions; paraventral areas densely covered by non-bracteate, hair-like extensions. Pectinate fringe of posterior margin hairs as on preceding segment.
Segment 4 with short acicular spine in middorsal position, not reaching the posterior margin of the segment ( Figs 12A View Fig , 14B View Fig ). Sensory spots present in subdorsal, midlateral and ventromedial positions ( Fig. 12 View Fig A–B); midlateral and ventromedial sensory spots considerably smaller than all other sensory spots in the species, but they occur consistently in all examined specimens. Glandular cell outlets type 1 present in subdorsal and ventromedial positions ( Figs 12 View Fig A–B, 14B–C). Pectinate fringe of posterior segment margin with long fringe tips from middorsal to ventromedial positions, and with very short tips between ventromedial positions. Pachycycli and cuticular hairs as on preceding segment.
Segment 5 with short acicular spine in middorsal position, not reaching the posterior margin of the segment ( Figs 12A View Fig , 14D View Fig ), and well-developed tubes in lateroventral positions ( Figs 12B View Fig , 14E View Fig ). Sensory spots present in subdorsal, midlateral and ventromedial positions ( Fig. 12 View Fig A–B). Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 6 with short acicular spines in middorsal and lateroventral positions ( Figs 12 View Fig A–B, 14D– E, 14G–H), not reaching the posterior margin of the segment. Sensory spots present in paradorsal, subdorsal, midlateral and ventromedial positions ( Figs 12 View Fig A–B, 15G). Females with female papillae in ventromedial positions, close to and lateral to sensory spots ( Figs 12B View Fig , 14E View Fig , 15H View Fig ); openings of papillae resemble small glandular cell outlets type 2 ( Fig. 15H View Fig ), and intracuticular substructures each form a semicircle with a small protuberance in the curved part of the structure ( Fig. 14E View Fig ). Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 7 with short acicular spines in middorsal and lateroventral positions, barely reaching the posterior margin of the segment ( Figs 12 View Fig A–B, 14D–E, 15G–I). Females with female papillae as on segment 6, but with openings slightly more anterior and lateral to sensory spots ( Figs 12B View Fig , 14E View Fig , 15 View Fig H–I); the substructure of these papillae differs from those on preceding segment, and forms instead a small intracuticular tube ( Fig. 14E View Fig ). Sensory spots, glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 8 with short acicular spines in middorsal and lateroventral positions, barely or just reaching the posterior margin of the segment ( Figs 12 View Fig A–B, 14D–G, 15G, I). Minute glandular cell outlets type 2 present in midlateral positions, but very close to the laterodorsal areas ( Figs 12A View Fig , 14G View Fig , 15F View Fig ). Sensory spots present in paradorsal, subdorsal, midlateral (posterior to glandular cell outlets), ventrolateral positions ( Figs 12 View Fig A–B, 15F, I). Females with female papillae similar to those on segment 7, but with
openings slightly more anterior and closer to midventral line ( Figs 12B View Fig , 14E View Fig , 15I View Fig ). Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 9 with acicular spines in lateroventral positions, just reaching the posterior margin of the segment ( Figs 12B View Fig , 15F View Fig ). Sensory spots, glandular cell outlets type 1 and 2 as on preceding segment, but female papillae not present ( Figs 12 View Fig A–B, 14F–G, 15F). Small, rounded nephridial sieve plates present in lateral accessory positions. Pectinate fringe of posterior segment margin with uniformly long fringe tips around the segment. Pachycycli and cuticular hairs as on preceding segment.
Segment 10 with well-developed laterodorsal tubes near posterior segment margin ( Figs 12 View Fig , 14J View Fig , 15 View Fig J–L). Sensory spots present in subdorsal (but close to paradorsal) positions ( Figs 12A, C View Fig , 15J, L View Fig ); males furthermore with sensory spots in ventrolateral positions ( Fig. 12D View Fig ). Glandular cell outlets type 1 present as two longitudinally arranged middorsal ones and in ventromedial positions ( Figs 12 View Fig , 14I View Fig ). The posterior segment margin of the tergal plate is straight, whereas margins of sternal plates are concave and extend midventrally into a point that almost reaches the posterior margin of the terminal segment; fringe tips of pectinate fringe are considerably shorter than those on preceding segments. Pachycycli and cuticular hairs as on preceding segment.
Segment 11 with lateral terminal spines ( Figs 11A View Fig , 12 View Fig A–B, 14A). Males with three pairs of penile spines ( Figs 12 View Fig C–D, 14J, 15K–L); dorsal and ventral penile spines are thin, flexible tubes, whereas the median ones are slightly thicker, conical, and more rigid; females with short, thin lateral terminal accessory spines ( Figs 12 View Fig A–B, 14I, 15J). Two pairs of sensory spots present in subdorsal positions and one pair in ventrolateral positions; one pair of subdorsal sensory spots medially on segment, other pair at posterior margin ( Fig. 15J View Fig ). Glandular cell outlets type 1 present as two longitudinally arranged middorsal ones and in subdorsal positions, anterior on segment. The dorsal and most of the ventral side of the segment are densely covered with non-bracteate hair-like extensions. Tergal extensions are wellspaced, short and pointed ( Figs 12 View Fig , 14 View Fig I–J, 15J, L); sternal extensions short, with ventrolateral seta-like tuft of extended fringe tips ( Figs 14J View Fig , 15J View Fig ).
Notes on habitat and ecology
The specimens collected in Kaneohe Bay, Hawaii, showed a very clear preference for muddy localities, rich in organic matter. They seemed to thrive very well in it, and appeared in high numbers. Especially at the locality in He’ia Fish Pond (St. MVS-0229-HI-06) – an 800 year old fish pond established by isolating a part of the coastal water with a 2 km long lava rock wall – specimens of E. sensibilis occurred in ball-like aggregations composed of detritus, and up to 40 specimens. At low magnification, these balls would just resemble detritus pellets, but by gently opening the ball with a needle, it would appear that the balls mainly consisted of mass aggregated kinorhynchs.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |