
Scinax ushiniauae, Gagliardi-Urrutia & Araujo-Vieira & Padial & Simões & Faivovich & Castroviejo-Fisher, 2024
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5406.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:9D3BB77F-2C7B-421C-B893-DDE8FC76F64A |
|
DOI |
https://doi.org/10.5281/zenodo.10630500 |
|
persistent identifier |
https://treatment.plazi.org/id/03CA87A8-FFFC-FFB7-EBC0-074C3492FE6A |
|
treatment provided by |
Plazi |
|
scientific name |
Scinax ushiniauae |
| status |
sp. nov. |
Scinax ushiniauae sp. nov.
( Fig. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Scinax sp. 18— Araujo-Vieira et al. 2023. Phylogenetic analysis and recognition as confirmed candidate species.
Holotype. CRBIIAP 2912 ( GGU 5494 ). Adult male collected on 5 May 2017 by Giussepe Gagliardi-Urrutia , Ehiko Rios-Alva , and Samantha Solís-Ruiz at Centro de Investigaciones Jenaro Herrera ( CIJH), Requena, Loreto, Peru (datum WGS84 ; 4.901539° S, 73.633465° W, 123 m a.s.l.). GoogleMaps
Paratypes. Twelve adult males and one female. CRBIIAP 1570, 1595, 1597, 1656, 2913 ( GGU 5313 , 5353 , 5358 , 5468 , 7618 ), MACN 59716 View Materials ( GGU 7617 ), MCP 14562 View Materials , 14564 View Materials ( GGU 5355 , 5586 ) , males, and CRBIIAP 1712 ( GGU 5578 ), female, collected on different dates in April and May 2017 and March 2022 by Giussepe Gagliardi-Urrutia , Ehiko Rios-Alva , José M. Padial , Katyuscia Araujo-Vieira, and Samantha Solís-Ruiz at the same locality as the holotype; CRBIIAP 1851, 1905, MCP 14565 View Materials , 15108 View Materials ( GGU 5099 , 6305 , 6301 , 5117 ) GoogleMaps , males, collected on different dates in March 2017 and April 2018 by Giussepe Gagliardi-Urrutia, Ehiko Rios-Alva, Samantha Solís-Ruiz, and Layné Guerra-Vargas at Río Blanco (RB), Frontera Village , Soplín, Requena, Loreto, Peru (datum WGS84 ; 5.88903° S, 73.76537° W, 131 m a.s.l.) GoogleMaps .
Referred specimens. MCP 14561 View Materials ( GGU 5350 ), juvenile, collected on April 2017 by Giussepe Gagliardi-Urrutia, José M. Padial, Ehiko Rios-Alva, and Samantha Solís-Ruiz at the same locality as the holotype; CRBIIAP 1883 ( GGU 5152 ), newly metamorphosed, collected in April 2017 by Giussepe Gagliardi-Urrutia , Ehiko Rios-Alva , and Samantha Solís-Ruiz at Río Blanco , Frontera Village , Soplín, Requena, Loreto, Peru GoogleMaps .
Diagnosis. (1) male SVL 20.2–22.9 mm; (2) nostrils not protruding dorsolaterally; (3) canthus rostralis poorly defined, straight or slightly convex; (4) loreal region slightly concave; (5) vocal sac weakly bilobate, ventrally not reaching the pectoral region; (6) dorsum uniform light to dark brown with scattered darker blotches of different sizes; (7) iris with median horizontal red streak, in life; (8) physiological chlorosis present, bones bluish-green; (9) moderately developed pre- and postaxial webbing of Toe IV, reaching the proximal half of the penultimate phalanx; (10) m. depressor mandibulae originating from the dorsal fascia that covers the suprascapula; and (11) advertisement call composed of a single pulsed note with duration of 4–62 ms, with 1–3 pulses emitted at a rate of 45.3–69.0 pulses/s.
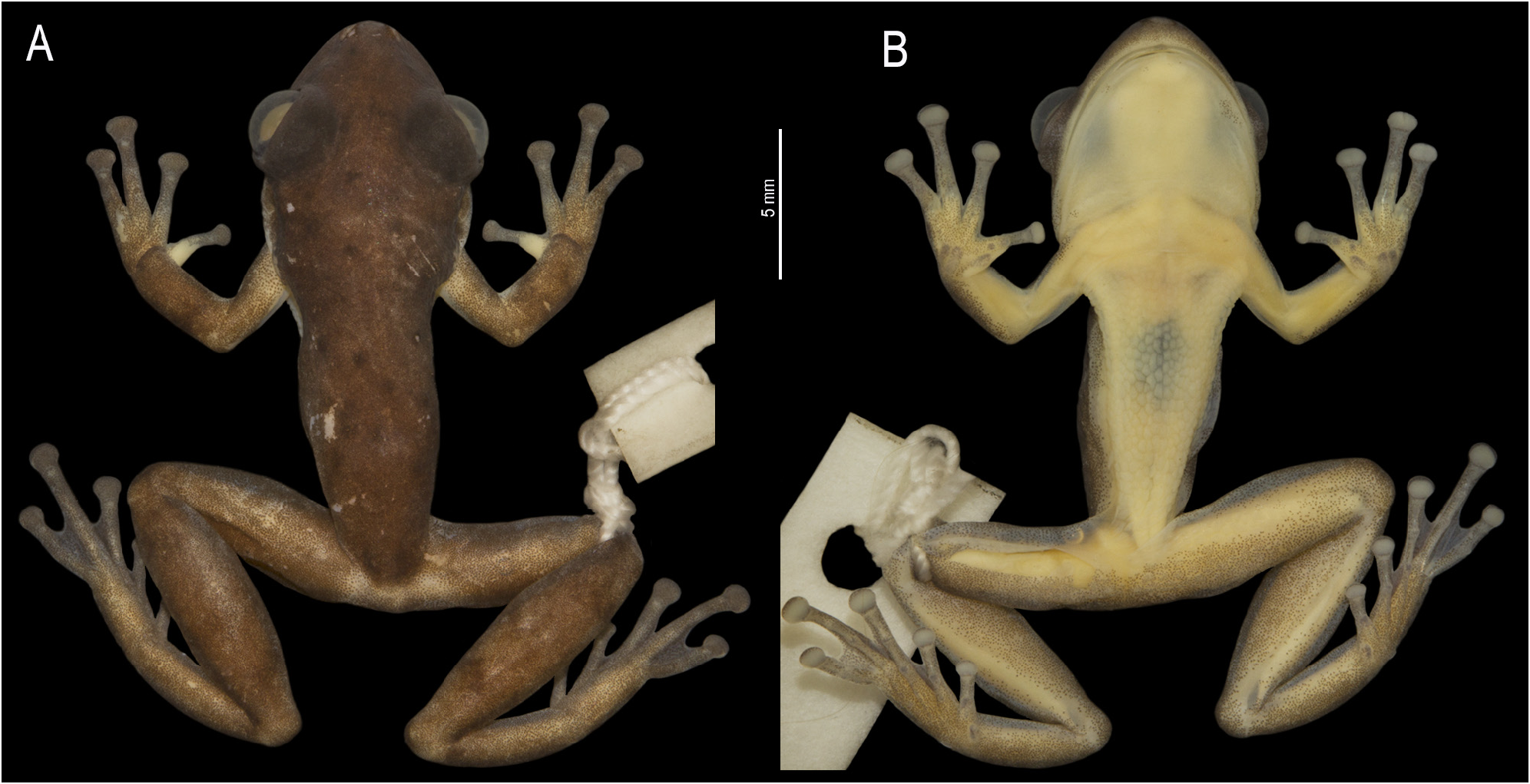
Description of the holotype. Adult male, 20.6 mm SVL ( Fig. 2A View FIGURE 2 ). Head longer than wide, HW 89% HL; HL 39% SVL; HW 35% SVL. Snout rounded in dorsal and lateral views; END equal to 76% ED; nostrils elliptical, not protruding dorsolaterally; the region between nostrils slightly concave; canthus rostralis poorly defined and slightly convex; loreal region slightly concave. ED 46% HW and 41% HL. Pupil horizontal. Palpebral membrane translucent, not reticulated. Tympanum distinct, round, small, TD 39% ED; tympanic annulus rounded, with upper portion hidden by the supratympanic fold. Supratympanic fold evident from the upper part of the tympanum to close to the insertion of the arm. Tongue ovoid, free laterally and posteriorly, shallowly notched posteriorly. Vomerine teeth in two barely separated transverse series, each bearing six (right) and five (left) teeth. Choanae oval. Vocal slits present, extending from the lateral base of tongue to the corner of the mouth. Vocal sac subgular, median, externally expanded, ventrally not reaching the pectoral region and not occupying space between head and body, with a slight medial constriction in the posterior portion ( Fig. 2B View FIGURE 2 ), which gives it a bilobate shape when deflated and inflated. Pectoral fold present, with pre-and postaxillar elements. Axillary membrane absent.
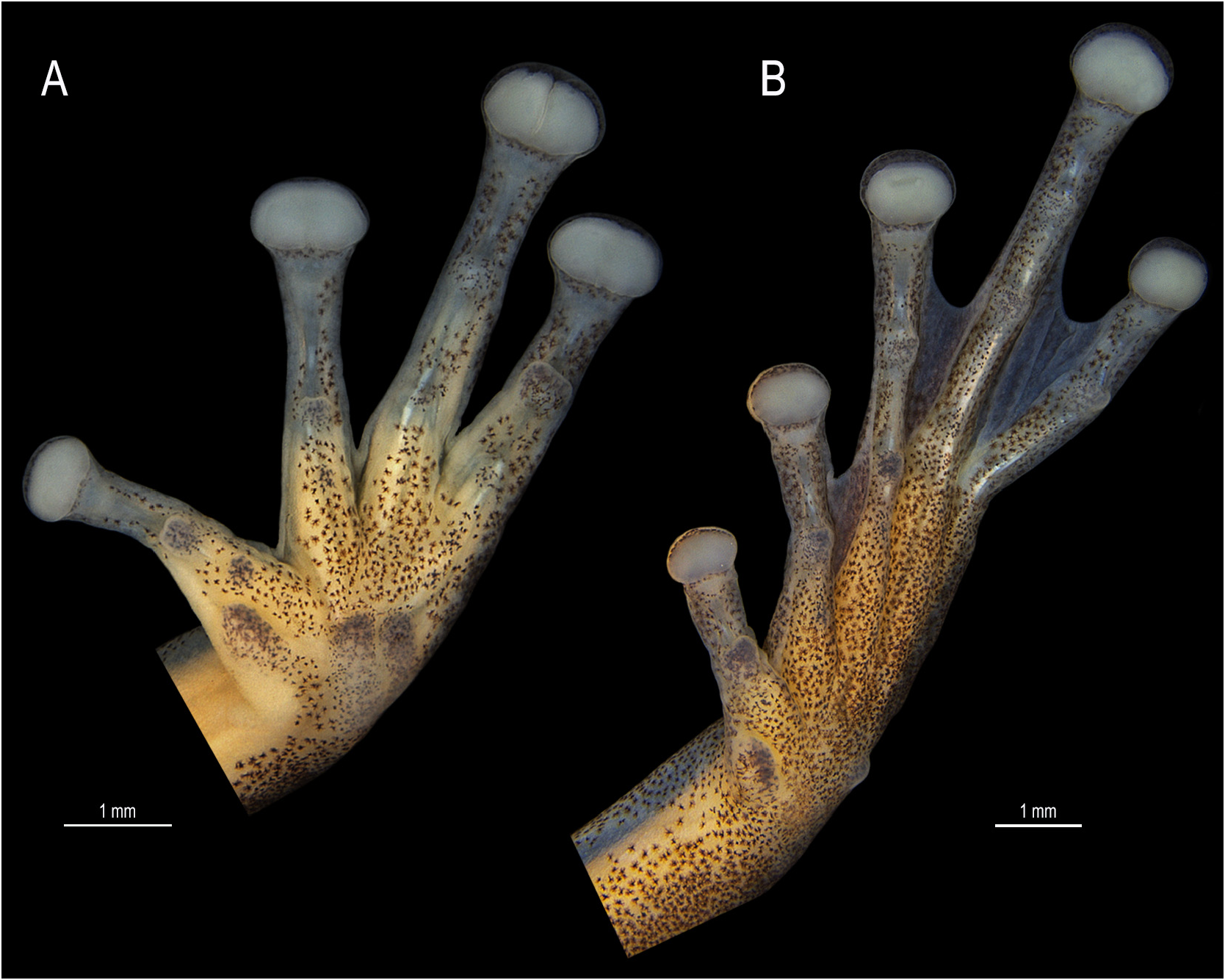
Forelimbs not hypertrophied; upper arm thinner than forearm. Ulnar tubercles absent. Fingers fringed; relative finger length II<III<V<IV ( Fig. 3A View FIGURE 3 ). Discs elliptical, wider than long, 3FD 33% ED and 85% TD; disc of Finger II smaller than others. Subarticular tubercles single, conical on fingers II and III, rounded and quadrangular on fingers IV and V, respectively. Supernumerary tubercles small, single, and rounded. Inner metacarpal tubercle single, flattened, and elliptical; outer metacarpal tubercle bifid and flattened. Basal webbing between fingers, more reduced between fingers II and III. Nuptial pad present on Finger II, thickened, light-colored, covering the metacarpal II dorsomedially, extending from the base of the inner metacarpal tubercle, obscuring its proximal and outer margins to distal margin of the subarticular tubercle ventrally, with distinct epidermal projections. Nuptial pad of finger III covers the metacarpal III dorsomedially, without epidermal projections.
Hind limbs proportionally long, TL 50% SVL, THL 45% SVL, FL 41% SVL. Tarsal fold and tubercles absent. Tubercles absent on heel. Toes fringed; relative toe length I<II<III≈V<IV ( Fig. 3B View FIGURE 3 ). Discs elliptical, wider than long, 4TD 27% ED and 69% TD. Subarticular tubercles single, conical on Toe I, and rounded on toes II–V. Supernumerary tubercles small, single, and rounded. Inner metatarsal tubercle single and elliptical; outer metatarsal tubercle single and elliptical (right toe) or round (left toe), three times smaller than the inner metatarsal tubercle. Webbing formula I 2 + – 21/4 II 11/2 – 21/2 III 11/2 – 21/3 IV 22/3 –1 + V ( Fig. 3B View FIGURE 3 ). Cloacal opening directed posteriorly at upper level of thighs.
Skin texture on dorsal surface of head, loreal region, upper eyelid, and tympanum region shagreen; small, flat, and scattered tubercles present on dorsal surface of trunk and flanks. Skin smooth on dorsal surface of limbs, gular and inguinal regions, and on hidden and ventral surfaces of forelimbs, tibiae, and tarsi. Low granules on abdomen, ventral surface of thighs, and subcloacal region. Glandular concentrations or spicule-shaped papillary epidermal projections absent on inner margin of forelimbs, pectoral and inguinal regions.
Measurements of the holotype (in mm). SVL 20.6; HL 8.1; HW 7.2; IND 1.9; IOD 2.3; ED 3.3; EN 2.5; TD 1.3; TL 10.4; FL 9.7; 3FD 1.1; 4TD 0.9.
Color in life of the holotype. Dorsum golden light brown with scattered black or dark brown blotches of different sizes ( Fig. 4A, B View FIGURE 4 ). Canthus rostralis with a diffuse black line, incomplete towards the nostril. Upper lip light brown or cream, with diffuse dark brown blotches below the eyes. Dorsal surfaces of forelimbs and hindlimbs conspicuously spotted with irregular transverse dark brown markings on forearms and dark brown blotches on shanks. Supracloacal region and knees with a light golden blotch dorsally. Flanks light yellow with a dark brown lateral irregular line from the posterior corner of the eye and through the upper part of the tympanic membrane and supratympanic fold to the inguinal region. Hidden surfaces of thighs, shanks, and inguinal region uniformly brown. Ventrally, fore- and hindlimbs light green. Soles and legs conspicuously spotted with brown. Palms, margins of gular region, area around arm insertion, and lateral edges of arms, thighs, and shanks finely spotted with brown. Pectoral region and abdomen white, immaculate; gular region yellow ( Fig. 4C View FIGURE 4 ). Iris silvery bronze with black reticulations and median horizontal red streak; red coloration also bordering the pupil; black blotches on the sides of the pupil. Physiological chlorosis present (bluish-green coloration in bones and lymph).
Color in preservative of the holotype. The coloration pattern is like that of the specimen in life, but darker. Golden tones faded and disappeared. Yellow, light green, and white colors became light beige or cream white. The iris coloration became black.
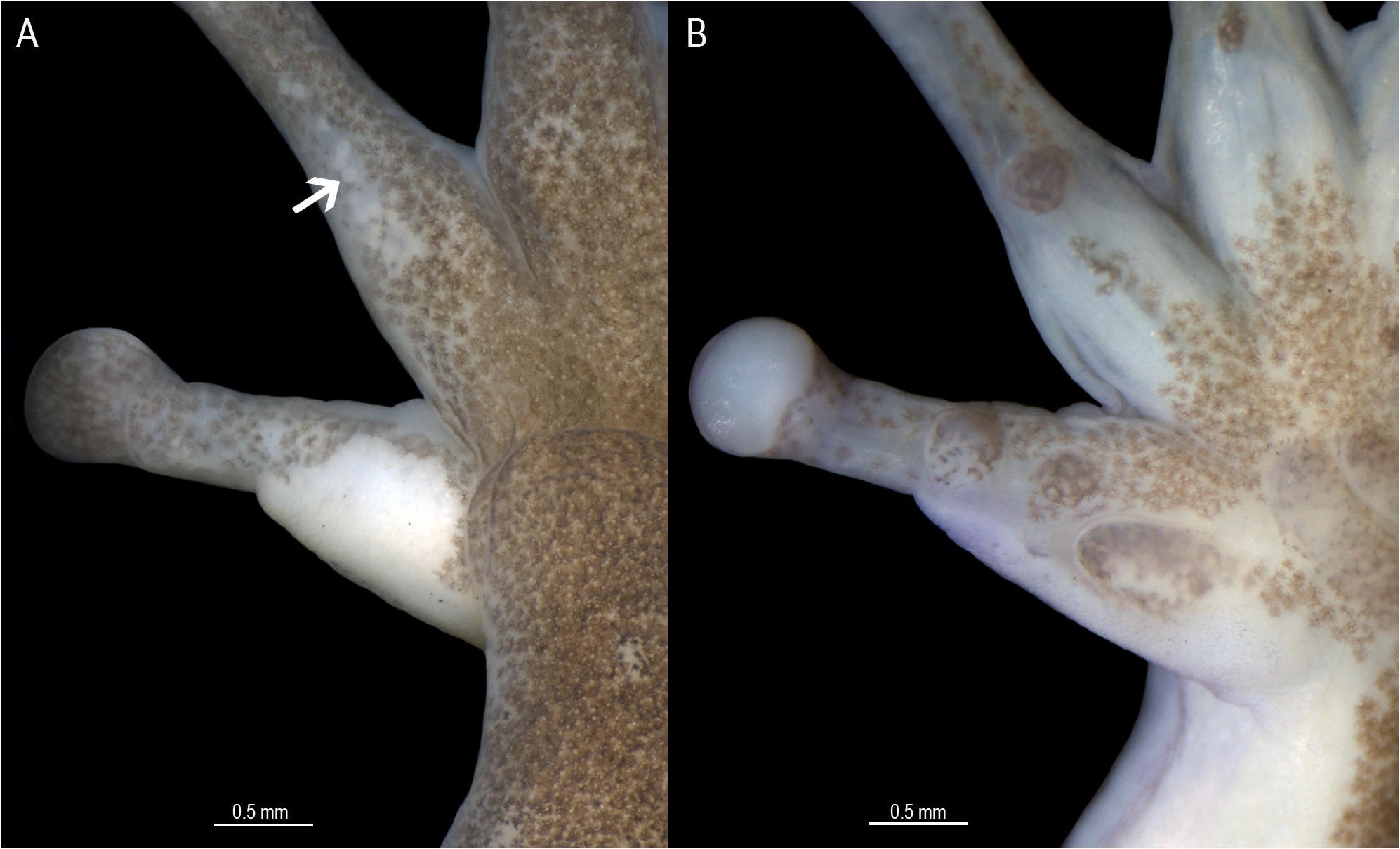
Variation in the type series. Some measurements are presented in Table 1 View TABLE 1 . Vomerine teeth in two transverse series barely separated or separated by a space equivalent to half the length of the dentigerous process. Vomerine teeth 3–5 on the right side and 2–7 on the left side. Nuptial pad on finger II like that of the holotype in seven specimens ( Fig. 5A, B View FIGURE 5 ), thinner in two specimens (CRBIIAP 1656, MCP 14564), and covering approximately half of metacarpal II dorsomedial surface in three specimens (CRBIIAP 1595, 1556, MCP 14562). Toe webbing formula varies as follows: I 2 + – (21/3 – 21/4) II (11/2 –1 +) – (21/2 – 21/4) III (11/2 –1 +) – (2 + – 2) IV (21/4 –2 +) – (11/3 –1) V. Skin texture on dorsum smooth or shagreen; smooth or with small, flat, and scattered tubercles on trunk and flanks; smooth or areolate on vocal sac and pectoral regions. The only available mature female (CRBIIAP 1712) is larger than males (27.3 mm versus 20.2–22.9 mm in males) and has webbing toe formula I 2 + – 21/4 II 1–2 + III 1–2 + IV 2 + –1 V. The horizontal pupil contracts extensively along the predominant (horizontal) and non predominant axis (vertical), generating in the most extreme contraction that we have documented ( Fig. 6F View FIGURE 6 ) a pupil that retains the horizontal shape, with an evident inflexion point along its ventral margin.
In life, dorsal color golden light brown, brownish gray to dark brown, with scattered or conspicuously distributed dark brown or black blotches of different sizes (see Fig. 6 View FIGURE 6 ); small light blotches are present on dorsum in most individuals (e.g., CRBIIAP 1851, 1570, 1597, 2905, MCP 14562; Fig. 6E View FIGURE 6 ). Canthal black line present (e.g., CRBIIAP 1597, 1656; Fig. 6C, E View FIGURE 6 ), absent (e.g., CRBIIAP 1570, MCP 14562; Fig. 6A View FIGURE 6 ), or indistinct (e.g., CRBIIAP 1595; Fig. 6B View FIGURE 6 ); when present, it covers the canthus rostralis completely (e.g., CRBIIAP 1587, MACN 59716; Fig. 6E View FIGURE 6 ) or just a small portion of it (e.g., CRBIIAP 1656; Fig. 6C View FIGURE 6 ). Number of transverse dark brown markings on forearms (e.g., 2–3 markings) and blotches on shanks (few or many blotches) variable among individuals; transversal dark brown markings on thighs present in some individuals (e.g., CRBIIAP 1597, MCP 14565; Fig. 6E View FIGURE 6 ). Flanks light or dark brown with complete (e.g., CRBIIAP 1570; Fig. 6A View FIGURE 6 ) or interrupted (e.g., CRBIIAP 2913; Fig. 6F View FIGURE 6 ), lateral, dark brown to black irregular line, extending from the posterior corner of the eye to the upper part of the tympanic membrane and supratympanic fold (e.g., CRBIIAP 1851, MCP 15108), to the middle of the flanks (e.g., CRBIIAP 1595, 1712, 1905; Fig. 6B, D View FIGURE 6 ), or the groin (e.g., CRBIIAP 1570, 1656; Fig. 6A, C View FIGURE 6 ). Ventrally, fore- and hindlimbs and gular region more conspicuously spotted with brown in MCP 14565; abdomen finely pigmented with dark brown in some individuals (e.g., CRBIIAP 1656, 1712). Iris silvery bronze or bronze, with finely or conspicuously distributed black reticulations. Gular region cream to white, finely spotted with brown on the mental area in the female (CRBIIAP 1712).
In preservative, coloration patterns are similar, but some specimens became darker (CRBIIAP 1712; Figs. 6D View FIGURE 6 , 7 View FIGURE 7 ) or lighter (CRBIIAP 2913; Figs. 6F View FIGURE 6 , 7 View FIGURE 7 ) after fixation in 10% formalin. Dorsally, dark and white blotches on dorsum disappeared (CRBIIAP 1597; Figs. 6E View FIGURE 6 , 7A View FIGURE 7 ) or became indistinct (CRBIIAP 1712; Figs. 6D View FIGURE 6 , 7A View FIGURE 7 ) in some specimens. Lateral lines on flanks faded and disappeared in the area immediately posterior to tympanum in some specimens (CRBIIAP 1597, 1851, 1905, MCP 15108). Two individuals (CRBIIAP 1851, MCP 15108) turned darker in some regions (head, nostril, and sacral region) after fixation.
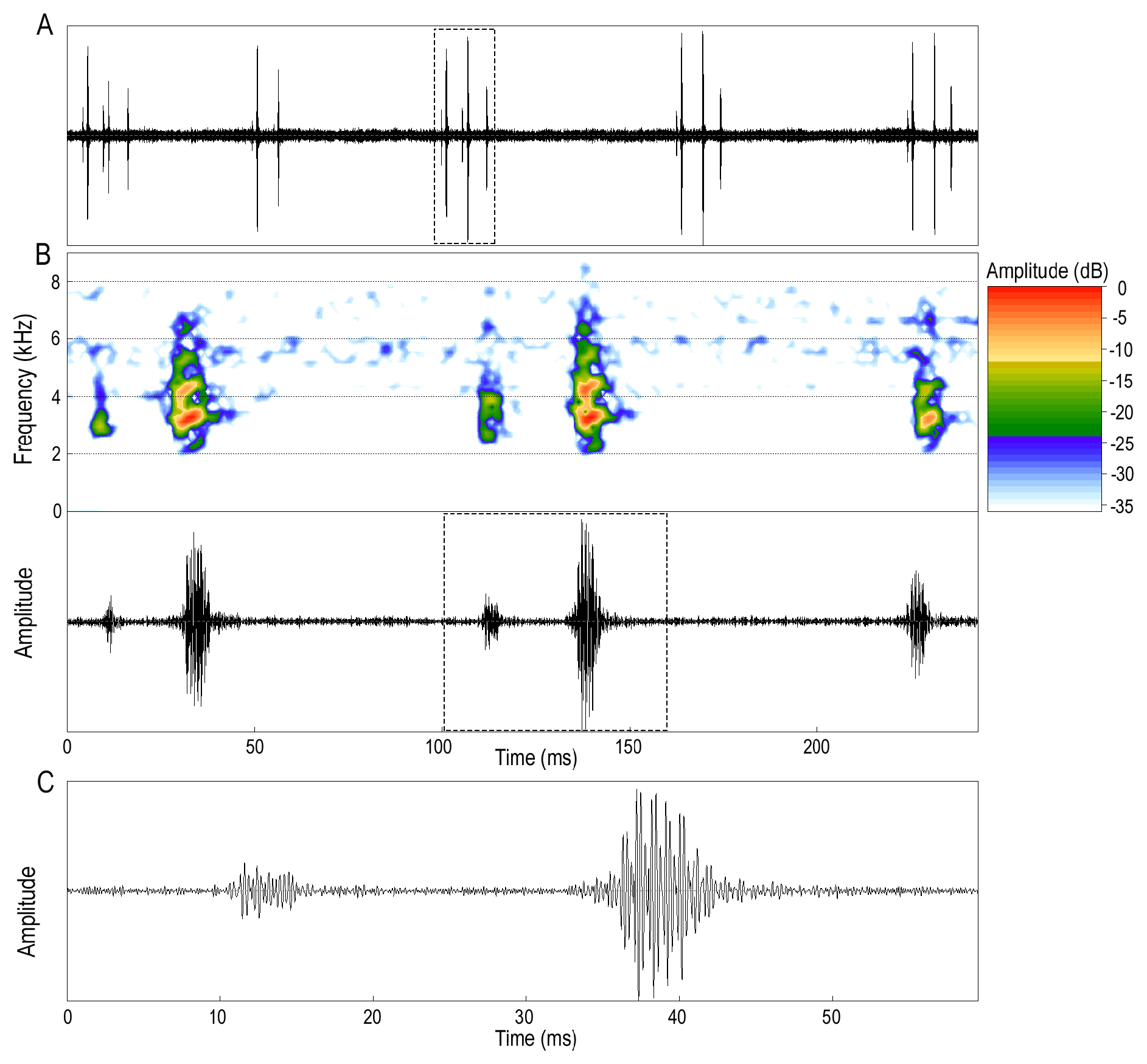
Advertisement call (mean ± SD in parentheses). The advertisement call of S. ushiniauae consists of a single pulsed note, with pulses of high amplitude preceded by pulses with lower energy ( Fig. 8 View FIGURE 8 ). Calls are emitted in series (n= 79 out of 88 analyzed calls) or as isolated notes (n = 9). Series are composed of 2–5 calls per series (2.6±0.2) with a call interval of 70–160 ms (60±0) and a duration of 90–380 ms (200±6). The individual call duration is 4–62 ms (29±14), and each is composed of 1–3 pulses (1.7±0.4) of 1–33 ms (9±2). In cases in which calls include two or more pulses (44 out of 88), the pulse rate is 45.3–69.0 pulses/s (58.2±6.7) with a pulse interval of 10–210 s (20±0). The dominant frequency is 2,713 –4,219 Hz (3,382±238), the 95% frequency 3,704 –6,460 Hz (4729±516), and the 25% frequency 2,627 –3,141 Hz (2,887±29).
Comparisons. Based on the phylogenetic hypothesis of Araujo-Vieira et al. (2023), we restrict our comparisons to the species of the S. cruentomma group. The smaller SVL (in mm) in males (20.2–22.9, n = 13) distinguishes the new species from S. blairi (male SVL 27.8–30.1, n = 13; Fouquette & Pyburn 1972) and S. karenanneae (male SVL 26.6–28.9, n = 2; Pyburn 1993).
The weakly bilobate vocal sac, not reaching the pectoral region, distinguishes the new species from S. altae , S. baumgardneri , S. blairi , S. cruentomma , S. exiguus , S. manriquei , S. staufferi (single vocal sac, reaching the pectoral region; Rivero 1961; Duellman 1972, 1986, 2001; Fouquette & Pyburn 1972; Barrio-Amorós et al. 2004; Araujo-Vieira et al. 2023).
The dorsal color pattern of the body, uniformly light to dark brown with scattered darker blotches of different sizes, distinguishes S. ushiniauae from S. albertinae , S. altae , S. exiguus , S. staufferi , and S. wandae (dorsum uniformly brown with longitudinal dark stripes; Duellman 1986, 2001; Pyburn & Fouquette 1971; Ferrão et al. 2022). The presence of a median horizontal red streak in the iris in living specimens distinguishes S. ushiniauae from S. albertinae , S. exiguus (median horizontal reddish brown streak in the iris; Duellman 1986; Ferrão et al. 2022), S. altae , S. baumgardneri , S. blairi , S. manriquei , S. karenanneae , S. lindsayi , and S. staufferi (median horizontal red streak in the iris absent; Rivero 1961; Duellman 2001; Fouquette & Pyburn 1972; Pyburn 1992, 1993; Barrio-Amorós et al. 2004). Furthermore, physiological chlorosis (impregnation of lymph, bones and some other tissues with biliverdin associated with its carrying serpin; Barrio 1965; Taboada et al. 2020) distinguishes the new species from S. altae and S. staufferi (chlorosis absent in these species; Duellman 1970).
The extension of pre- and postaxial webbings of Toe IV reaching the proximal half of the penultimate phalanx distinguishes the new species from Scinax albertinae , S. exiguus , and S. wandae (pre- and postaxial webbings of Toe IV not surpassing the proximal half of antepenultimate phalanx anteriorly; Pyburn & Fouquette 1971; Duellman 1986; Ferrão et al. 2022). The m. depressor mandibulae with an origin from the dorsal fascia that covers the suprascapula differentiates Scinax ushiniauae from S. staufferi (m. depressor mandibulae without an origin from the dorsal fascia that covers the suprascapula; Faivovich 2002; Araujo-Vieira et al. 2023).
The advertisement call consisting of a single, very short pulsed note, characterized by the combination of note duration 4 to 62 ms, 1–3 pulses/note, and pulse rate of 45.3–69.0 pulses/s differentiates Scinax ushiniauae from all species of the S. cruentomma group whose calls have been described (see the Discussion section about the call of S. manriquei ; Barrio-Amorós et al. 2004): S. albertinae (note duration 500–650 ms, 79–105 pulses/note, and pulse rate of 111–200 pulses/s, n = 3; Ferrão et al. 2022: figs. 6A, B, 13A), S. altae (note duration 140–180 ms and 110–130 pulses/s, n = 7; León 1969: pl. 4C; Duellman 1970: table 17), S. baumgardneri (note duration 530–700 s and 170–180 pulses/s, n = 5; Duellman 1972: fig. 4A), S. blairi (note duration 140–160 ms and 18–22 pulses/note, n = 9; Fouquette & Pyburn 1972: fig. 3), S. cruentomma (note duration 220–370 ms, 39–54 pulses/note, and pulse rate of 153.8–192.8 pulses/s, n = 6; Duellman 1972: fig. 4B; Duellman & Pyles 1983: table 2; Carvalho et al. 2015: fig. 2), S. exiguus (note duration 200–1,700 ms, 23–90 pulses/note, and pulse rate of 64–125 pulses/s, n = 9; Duellman 1986: fig. 2C; Carvalho et al. 2017: fig. 2), S. lindsayi (note duration 80–100 ms, n = 3; Pyburn 1992: fig. 2); S. staufferi (note duration 130–230 ms and 100–130 pulses/s, n = 18; León 1969: pl. 4B; Duellman 1970; pl. 26, fig. 2), S. strussmannae (note duration 100–110 ms, 23–27 pulses/note, and pulse rate of 200–250 pulses/s, n = 1; Ferrão et al. 2018a: fig. 5), and S. wandae (note duration 600–710 ms, 70–108 pulses/note, and pulse rate about 130 pulses/s, n = 5; Pyburn & Fouquette, 1971: fig. 2; Pombal et al. 2011: fig. 7F).
Scinax ushiniauae morphologically resembles S. strussmannae ( Ferrão et al. 2018a: figs. 1–4), differing mainly in traits of their advertisement calls. However, the new species can be further differentiated from S. strussmannae by having nostrils not protruding dorsolaterally, canthus rostralis poorly defined, straight or slightly convex, and loreal region slightly concave (nostrils protruding dorsolaterally, canthus rostralis well defined, curved medially, and loreal region markedly concave in S. strussmannae ; Ferrão et al. 2018a).
Etymology. The specific name ushiniauae honors Meri del Pilar Ushiñahua Álvarez, a recently retired professor from the Faculty of Biological Sciences of the Universidad Nacional de la Amazonía Peruana—UNAP. During decades of teaching, she has stimulated students with her enthusiasm and joy, her passion for the conservation and management of Amazonia, and her studies of Amazonian turtles. Her academic legacy lives through the large number of students she trained throughout her academic career.
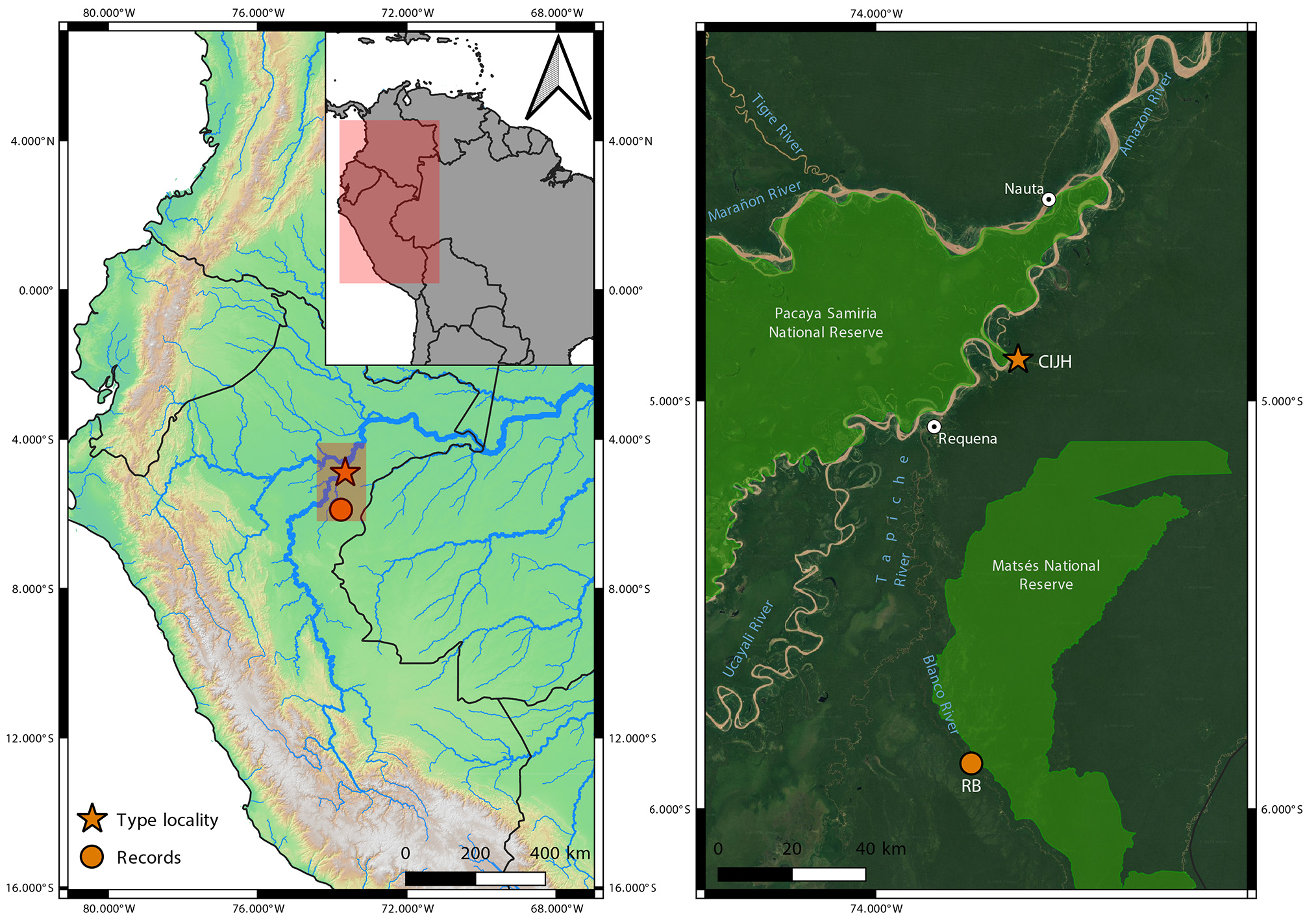
Geographic distribution and natural history. The new species was collected in two Peruvian localities, CIJH and RB, in the province of Requena, department of Loreto, covering an area of just over 100 km in a straight line, in which there are no large rivers that fragment the populations ( Fig. 1 View FIGURE 1 ). The CIJH comprises terra-firme (unflooded) forest environments growing on clay and white-sand soils and forests growing on hydromorphic, seasonally flooded soils. The rainy season spans approximately four to six months, with the heaviest rainfall from December to March. Mean annual precipitation is 2,521 mm, and the average yearly air temperature is 26 °C (22.5–31.9°C; López-Parodi & Freitas 1990; IIAP 2023). RB is located approximately 110 km south of CIJH, on the east bank of the TapicheBlanco interfluve. Part of this interfluve is constituted by large areas of white-sand forests and other vegetation types characteristic of white-sand soils (known locally as varillales and chamizales; Pitman et al. 2015; GarcíaVillacorta et al. 2016). Hydromorphic soils and terra-firme clay-rich soils are also common at this site, home to a megadiverse plant community ( Pitman et al. 2015). RB is also in the buffer zone of the Matsés National Reserve. A population that may belong to the new species occurs in the city of Leticia, Departament of Amazonas, Colombia (J. M. Padial, pers. obs.), although its identity remains uncertain.
Most individuals of S. ushiniauae (55 %) were found in white-sand forest environments, while the remaining 45 % were located in forests growing on clay-rich soils ( Gagliardi-Urrutia 2020). Males were found vocalizing in horizontal or vertical positions, with the head upwards, while perched on varillales and chamizales vegetation (0.7–1.5 m above ground) that grow on the margin or adjacent areas of temporary ponds located at the edges of old timber extraction trails inside the forest, or ponds in natural depressions in the forest, always associated with white-sand soils ( Fig. 9A, B View FIGURE 9 ). These ponds were formed when rainwater filled depressions, resulting in land subsidence processes. The new species shares the same reproductive habitats with Leptodactylus diedrus Heyer.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.