
Altica cyanea Weber, 1801
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3918.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:F87634FE-2F58-476A-9A9F-B31555B13041 |
|
DOI |
https://doi.org/10.5281/zenodo.5696588 |
|
persistent identifier |
https://treatment.plazi.org/id/03CB6450-FF91-F97A-76A8-CD8DFC67495A |
|
treatment provided by |
Plazi |
|
scientific name |
Altica cyanea Weber, 1801 |
| status |
|
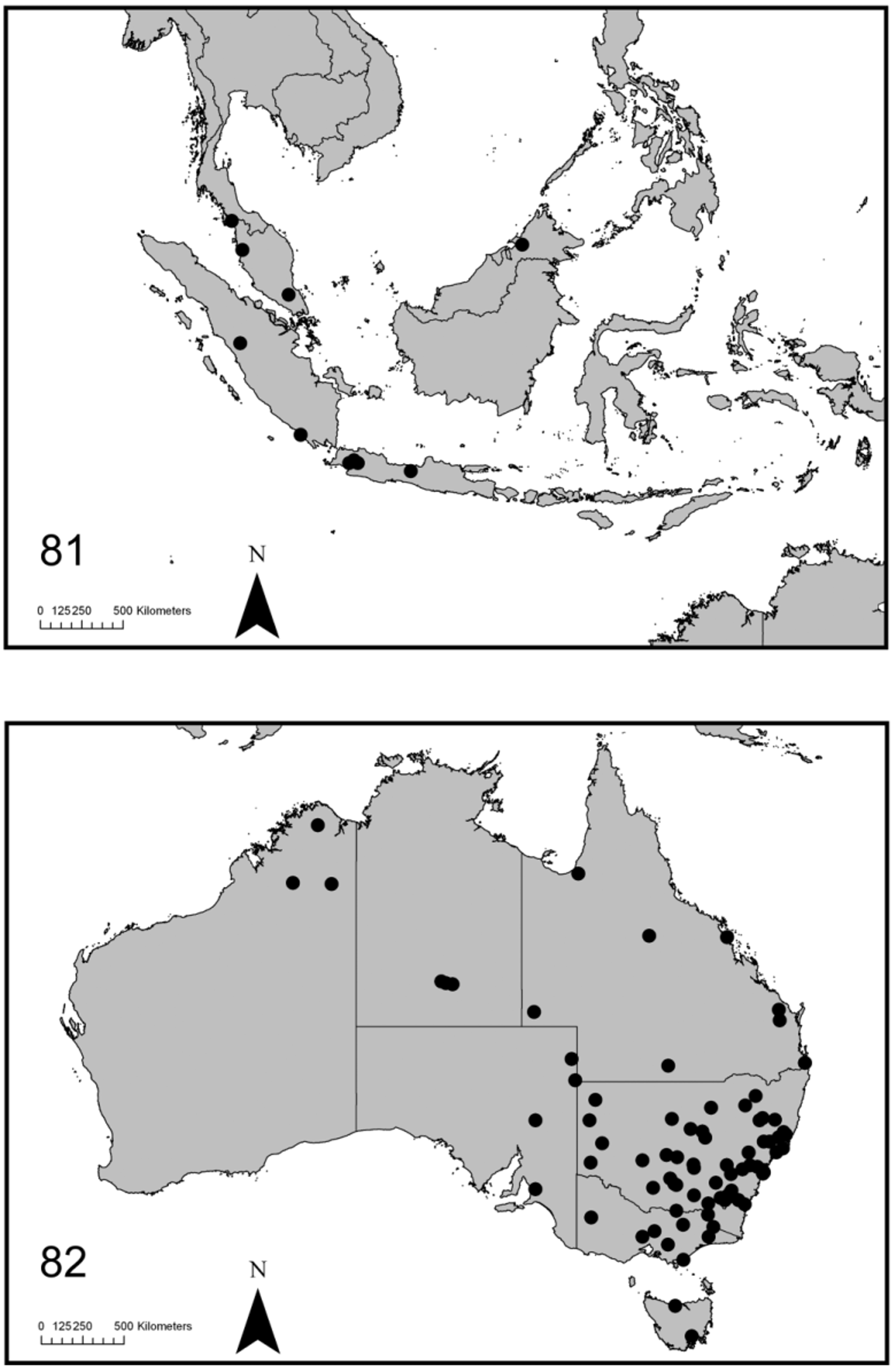
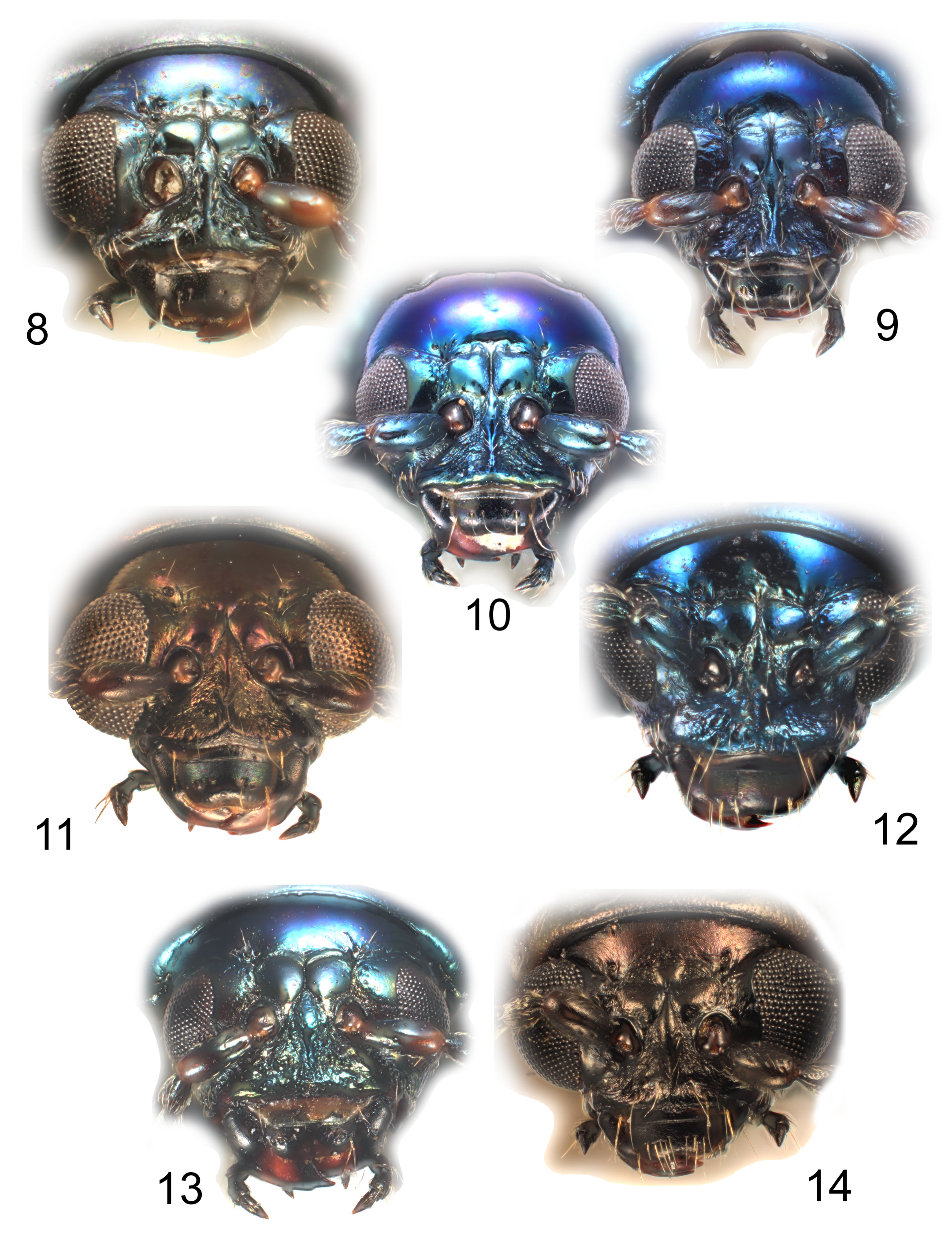
( Figs 6 View FIGURES 1 – 7 , 13 View FIGURES 8 – 14 , 28 View FIGURES 27 – 29 , 41, 42, 56, 57, 71, 72 View FIGURES 62 – 76 , 81 View FIGURE 81 )
Altica cyanea Weber 1801: 57 (type locality Sumatra).
Haltica cyanea: Illiger 1807: 115 .
Galleruca cyanea Fabricius 1801: 497 (objective junior homonym, type locality Sumatra). Haltica ianthina Illiger 1807: 115 (nom. nov. pro cyanea Fabricius 1801 , nec Weber 1801). Altica caerulea partim sensu auctt. nec Olivier, 1791; Gressitt & Kimoto 1963: 889.
Material examined (20, * = dissected specimen). Type material: Altica cyanea Weber : Lectotype (this designation): ♀*/ cyanea / ( NHMD).
Non-type material: Indonesia: Java: ♀*, ♀/ Buitenzorg, vi.1907 ( BMH); 2♂ */ Java, Dieng Plateau, above Telago Warna, 2150m, on low shrub, ii.1991, C Reid ( ANIC); ♂*, ♀*/ West Java E entrance to Gunung Halimun NP, ix.1999, C Reid ( AMS); Sumatra: 2♂ *, ♀/ West Lampung, 3k S Krui, Melastoma , old coastal palm plantn, 19.xii.2001, C Reid ( AMS); Malaysia: ♀*, 4/ North Borneo or Sarawak, 1958-9, TC Maa ( BMH); 1/ British North Borneo [Sabah], Tenompok, 48k E Jesselton, 1460m, 26–31.i.1959, TC Maa ( BMH); ♂*, ♂, ♀*, ♀, ditto, 19.ii.1959 ( BMH).
Description. Length: male 5.1–5.8mm, female 5.2–6.1mm.
Colour: dorsum usually deep blue, rarely purplish or bicoloured, either with green pronotum and blue elytra or the mostly blue elytra narrowly green laterally; base of head often reddish (teneral specimens?); first antennomere dark brown with red apical third and base; remainder of antennae black or antennomeres 2 & 3 reddish-brown; venter and legs entirely black, or legs partially reddish-brown (teneral specimens?), with duller metallic reflection than dorsum.
Head: ratios (Tables 3 & 4): male: EG 2.80–4.00; IE 1.57–1.82; HG 7.80–10.00; HN 1.00–1.08; NE 2.35–2.66; female: EG 3.40–4.00 ( A. cyanea lectotype: 3.40); IE 1.55–1.63 ( A. cyanea lectotype 1.62); HG 8.8–9.80 ( A. cyanea lectotype 8.8); HN 1.03–1.06 ( A. cyanea lectotype 1.04); NE 2.33–2.50 ( A. cyanea lectotype 2.50); pubescence: several small setae at posterior of orbit, longitudinal row of short recumbent setae at sides of frontoclypeal ridge, 1 long pair and several short setae behind clypeal anterior margin; transverse row of long setae between eye and buccal cavity; face impunctate except small punctures at bases of orbital setae; vertex without microreticulation; postantennal calli smooth, not microreticulate; eyes small and relatively flat; postantennal calli usually almost quadrate, with bluntly triangular anterior angles and truncate to convex bases, these usually tranversely subdivided, with secondary triangular areas; calli variably adjacent, from most of length to base only; frontoclypeal ridge lanceolate, broad and smooth or finely rugose at base, entirely convex, anteriorly terminating in a narrow keel before clypeal margin or keel absent; anterior edge of clypeus dull and finely rugose, strongly raised, often as a prominent transverse ridge, sides of clypeus microreticulate and finely rugose.
Thorax: pronotum with or without pair of shallow ovate depressions behind anterior border; shining, entirely non-microscuptured or apical half and basal field microreticulate; non-glandular puncturation variable, from small sparse punctures only laterally, to anterior and basal field with sparse (separated by at least 2 diameters) larger punctures, but disc always finely and sparsely punctured; hypomeron without microreticulation, smooth, except anterior angles finely transversely strigose and sometimes with sparse punctures; prosternum anteriorly smooth and shining, process finely transversely grooved and dull, slightly rugose; scutellum triangular with curved sides, to semiovate, microreticulate or shining and smooth; elytra shining, microreticulation absent or at extreme apices only, strongly and closely but irregularly punctured, interspaces mostly 0.5–2 diameters, sometimes with smooth elongate intervals on disc; elytra with small deep depression laterally behind humerus, but without distinct keel; femora densely microsculptured and pubescent; outer face mid tibia convex, usually with prominent keel for most of length, apical fifth sometimes flat; male first protarsomere distinctly longer than broad, female 1.6–1.7x longer than broad.
Abdomen: abdominal ventrites densely microsculptured and with recumbent pubescence; male: penis 1.95–2.15mm long; in dorsal view parallel-sided to the rounded apex, which has a protruding short truncate lobe; dorsal and ventral surfaces slightly curved in lateral view, with extreme tip recurved; shallowly transversely ridged on middle of dorsal surface (only 2 ridges on specimen from Dieng, Java); venter without transverse or oblique ridges, two long apicoventral depressions present, 0.30–0.35x penis length, ovate, internally finely ridged or smooth surfaced, laterally smooth edged, separated by a narrow sharp ridge; female: tignum 1.04–1.28mm long, basal part broad and irregularly outlined, lateral arms large and broadly triangular and apex broadly spatulate; spermathecal collum of variable length and thickness and containing 1–2 twists; vaginal palpi short and broad, almost ovate, length: width ratio 0.6–1.2, with rounded to truncate apex and straight or convex inner margin; palpal apodemes 3– 5 x length palpi, 0.5–0.5x width palpi.
Distribution and biology. Altica cyanea appears to be endemic to Sundaland (Peninsular Malaysia, Sumatra, Borneo and Java) and is absent from Australia and islands east of Java. Manfred Döberl has kindly provided additional records of A. cyanea based on his dissections and all fit this limited distribution on the Sunda shelf: Malaysia: Endau Rompin NP, Johore; Taiping, Perak; Indonesia: Cibodas & Cipanas, Java; Tebing Tinggi, North Sumatra; Payakumbuh, West Sumatra.
Altica cyanea has been found feeding on Melastoma (Melastomaceae) View in CoL in Sumatra and Java (pers. obs.; label data, AMS). Published records of hosts for probably correctly identified A. cyanea include only Melastoma View in CoL ( Kamarudin & Shah 1978; Ooi 1987). Mohammedsaid (2004) has also listed Melastoma View in CoL as the host of A. cyanea but his entry for this species is a conflation of A. cyanea and A. aenea . Melastoma malabathricum View in CoL is a weed in forestry plantations and A. cyanea has been proposed as a biocontrol agent for it ( Kamarudin & Shah 1978). Altica cyanea and its closest relative A. birmanensis have been collected together in Java (Dieng Plateau).
Taxonomic notes. Altica cyanea Weber, 1801 , was described from material collected by the Danish naturalist Daldorff in Sumatra. In the same year Fabricius named A. cyanea from the same material ( Fabricius 1801), without acknowledging Weber’s prior use of the name. Weber’s publication appeared at least by August and probably before March, 1801 ( Evenhuis 1997; Y. Bousquet, pers. comm. 2014), whereas Fabricius’ publication is not datable within 1801 and therefore should be dated 31 December 1801 (ICZN: Art. 21). Weber was Fabricius’ pupil and it is possible that pupil and master used the same name for the same insect without realising that each was going to publish it. Daldorff was a Danish Naval officer in the Danish East Indian Company, based in Tranquebar on the east coast of India ( Daldorff 1797), and is known to have visited Bencoolen [Bengkulu], the principle British trading port on the west coast of Sumatra in 1800 ( Heyne 1814). Therefore Daldorff’s material was probably from Bengkulu. Weber’s description was minimal: blue, but including a greenish variety, elytra punctate, similar and related to A. oleracea [from Europe] but more elongate, and with dense short grey pubescence on the venter. This description fits any of the three species of Altica View in CoL known from Sumatra in collections. Fabricius’ description was also minimal: blue, shining, antennae and legs black, similar to A. tamaricis [from Europe] but slightly larger and abdomen black. Illiger in Berlin also had material from Daldorff and based on comparison of Weber and Fabricius’s descriptions with his material decided that two species were involved. He therefore renamed Fabricius’ junior homonym, using his emendation of the genus name: Haltica ianthina Illiger, 1807 .
Weber’s early beetle types were retained in Fabricius’ collection, housed in NHMD ( Horn & Kahle 1937) and there should be at least two syntypes, since a colour variety was mentioned. However, Gressitt claimed to have examined the syntypes of A. cyanea in the Royal Scottish Museum (now NMSE) in Scotland in the 1950s ( Gressitt & Kimoto 1961, 1963). Scherer (1969) also listed RSM as repository of the types but without any evidence that he had seen the material ( Scherer 1969). A search of the NMSE collections has revealed that there are two syntypes of Altica caerulea Olivier, 1791 (q.v.), but no Weber or Daldorff material (Richard Lyszkowski, pers. comm. 2013).
We think Gressitt muddled his notes when travelling around collections in Europe, and this error was simply copied by Scherer. There is a single specimen in NHMD, labelled as a type of Galeruca cyanea Fabricius, 1801 , collected by Dahldorff in Sumatra and there is a single early specimen in ZMB standing as a type of Haltica ianthina Illiger, 1807 . The handwritten label on the first appears to be by Fabricius.
Documentation is lacking for the origins of these specimens in NHMD and ZMB, therefore at least three scenarios are possible: (i) these are the two syntypes of Weber, one relabelled by Fabricius and the other relabelled by Illiger; (ii) the specimen in Copenhagen is one of Weber’s syntypes, relabelled or also used by Fabricius, and the specimen in Berlin is not a type; (iii) Weber’s syntypes are missing and only Fabricius’ specimen remains. In the interests of reducing ambiguity for this species, we consider it reasonable to suppose that the same specimen was used by pupil (Weber) and master (Fabricius) and that this is the specimen in NHMD. Furthermore that the specimen in Berlin is from the same collection of material (by Daldorff) but has no type status. We designate the NHMD specimen a lectotype of Galeruca cyanea Weber, 1801 , to cement this argument.
Altica cyanea has been confused with both A. birmanensis and A. caerulea for most of the last 100 years. Maulik (1926) synonymised A. cyanea and A. birmanensis , followed by Scherer (1969) and Medvedev (2009), although the two last authors based their arguments on misidentifications. Kimoto separated the species on the presence or absence of a lateral elytral groove (and keel) without mentioning genitalic characters ( Kimoto 2000: 256). This character is useful but the genitalia are the only reliable diagnostic features. Altica cyanea is found wholly within the range of A. birmanensis , which suggests the possibility that it is a divergent population within that species. Further work is needed to determine the status of these species.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Galerucinae |
|
Genus |
Altica cyanea Weber, 1801
| Reid, C. A. M. & Beatson, M. 2015 |
Haltica cyanea:
| Illiger 1807: 115 |
Altica cyanea
| Weber 1801: 57 |
Galleruca cyanea
| Gressitt 1963: 889 |
| Illiger 1807: 115 |
| Fabricius 1801: 497 |